Гаплогруппы коренных древних европейцев,гаплогруппа I I1 и I2 у…
Если среди современных европейских народов Европы можно выделить несколько основных гаплогрупп (более 5%), это конечно же R1b, R1a, I1, N1, I2, G2 на севере и J, E1b1, T на юге Европы, то касательно населения древней Европы ситуация совсем иная.
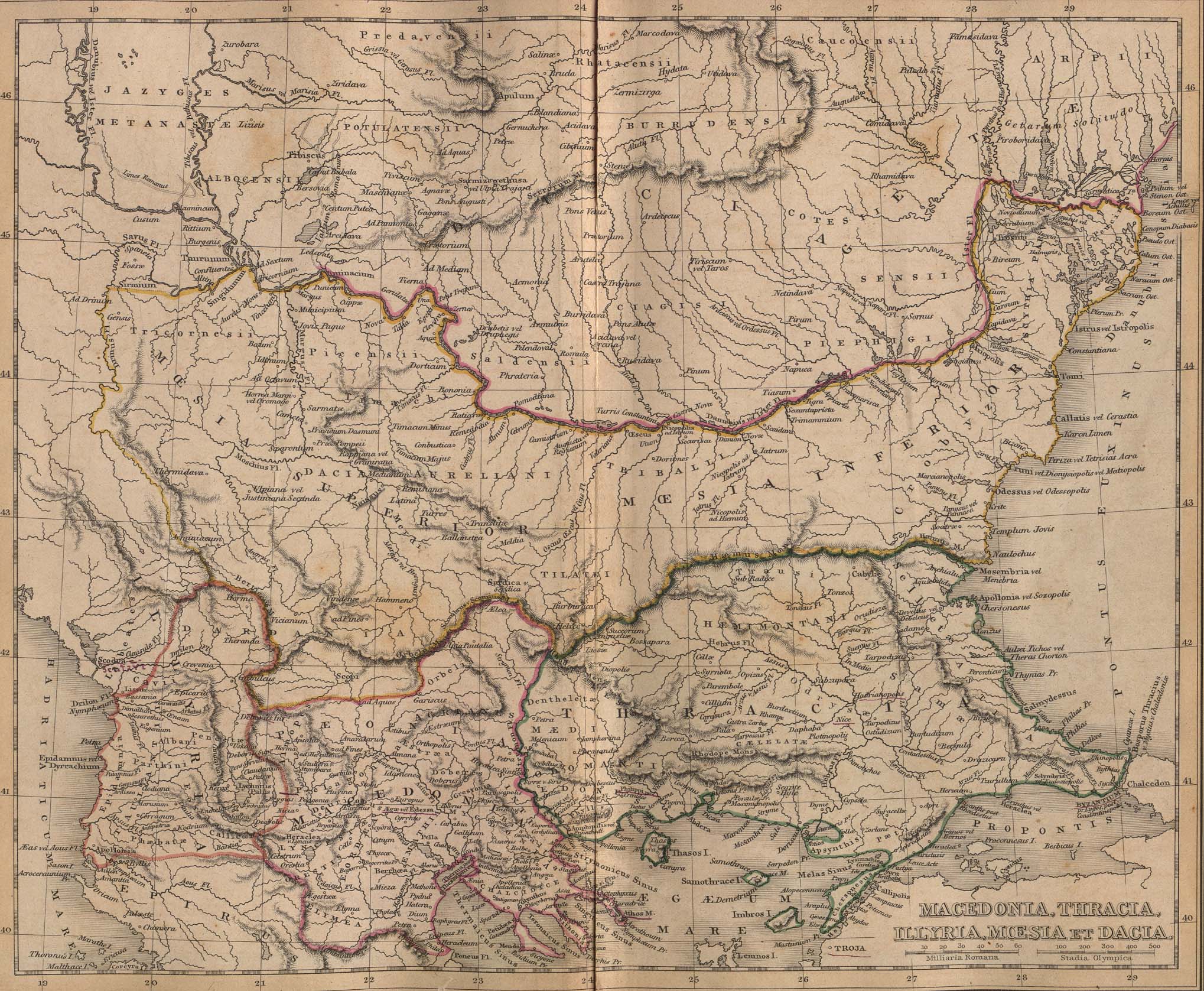
Убежища последнего ледникового максимума
Происхождение гаплогруппы I
Происхождение древних европейцев несомненно можно отождествлять с ДНК гаплогруппой I, отделившейся от пришедшей с Ближнего Востока Y гаплогруппой IJ, которые были кроманьонцами.В в популяционной генетике человека, Гаплогруппа I (M170, P19, M258) Y — хромосомная гаплогруппа, предположительно зародилась 20-25 тысяч лет назад на Балканском полуострове. Гаплогруппа I встречается почти у 1/5 европейского населения.
Некоторые исследователи полагают, что местом появления гаплогруппы I могла быть Граветтская культура, своего рода обитаемый "оазис", где люди переживали Последний ледниковый максимум, который произошел примерно 21 тысяч лет назад. Со времён прихода в Европу людей современного вида (кроманьонцев) около 40 тыс. лет назад и до индоевропейского вторжения около 5 тыс. лет назад, по всей видимости, гаплогруппа I составляла большинство населения Европы, возможно, существовало большее количество субклад, ныне исчезнувших, из которых ныне идентифицировано лишь две (I1 и I2). Строители западноевропейских мегалитов, вероятнее всего, принадлежали к гаплогруппе I2a.
Этногеографическое распространение гаплогруппы I
Наибольшее распространение гаплогруппы I имеется в 2 очагах распространения: первый это гаплогруппа I1 c центром в Скандинавских странах среди шведов, финнов, датчан и норвежцев.Второй это гаплогруппа I2 с центром на Балканах в Хорватии и Сербии.И гаплогруппа I1 и I2 распространены и в остальных странах Европы приблизительно в соотношении 15% в среднем.
Гаплогруппа I1
Гаплогруппа I1 является коренной европейской гаплогруппой. Большинство современных носителей гаплогруппы I1 составляют носители германских языков индоевропейской семьи, хотя первоначально данная гаплогруппа была связана, по-видимому, не с индоевропейскими народами, а с догерманским субстратом (Культура воронковидных кубков). I1 идентифицируется по меньшей мере 15 уникальными мутациями, что означает, что данная группа либо была совершенно изолирована в течение долгого периода (что маловероятно), либо пережила серьёзное «бутылочное горлышко» в сравнительно недавнем времени. Хотя первая мутация, отделившая I1 от I, могла произойти ещё 20 тысячелетий назад, все сегодняшние носители этой ДНК гаплогруппы происходят от одного мужчины, жившего не раньше, чем 5 тысячелетий назад. Это вполне совпадает со временем прихода на Скандинавию индоевропейцев, которые, как предполагается, уничтожали большую часть мужчин коренного населения или ставили их семьи в невыгодное демографическое положение. Так что выглядит вполне правдоподобным, что это вторжение пережил лишь один род коренных скандинавов (или, например, возможно, один мальчик), потомки которого в дальнейшем и составили гаплогруппу I1, которая таким образом стала достоверной меткой скандинавского протогерманского этноса, складывавшегося в ту эпоху. Сегодня представителей этой группы находят всюду, куда имели место вторжения или миграции древних германцев, единственным исключением может быть назван север европейской части России (Карелия, Вологда) и большая часть Поволжья, куда гаплогруппа I1 передалась через финнов.
Гаплогруппа I2
Гаплогруппа I2 имеет 3 основных субклада с разным этногеографическим распространением в Европе:
Субклад I2-M26 это одна из основных Y-хромосомных линий на Сардинии (достигает 40%) и у басков. С низкой частотой встречается также по всей юго-западной Европе: В Испании, Франции, Португалии, Италии.Субклад I2-M423 характерен для населения Балкан и Карпат, чаще всего наблюдается у динарийских славян (сербы, хорваты и боснийцы), а также в Молдове и на юго-западной Украине. Также с заметной частотой обнаруживается в Албании, северной Греции, Болгарии, Словакии, на восточной Украине, в Белоруссии и юго-западной России.Субклад I2-M223 характерен для северо-западной Европы и достигает максимума в Германии (Нижняя Саксония). Также встречается в Румынии, Молдавии и с низкой частотой в Восточной Европе.
Гаплогруппы современных Европейцев после индоевропейской экспансии и Великого переселения народов
Среди современных этнических европейцев распространены следующие гаплогруппы в порядке убывания (включая южную и восточную Европу): гаплогруппа R1b1a2 (33.71%),гаплогруппа R1a1 (22.21%), гаплогруппа I1 (8.39%), гаплогруппа I2a (7.97%), гаплогруппа E1b1b1 (6.77%),гаплогруппа J2 (6.48%), гаплогруппа N1c1 (5.82%), гаплогруппа G2a3 (3.92%), гаплогруппа I2b (2.20%) и с низкой частотой встречаются гаплогруппы T (1.82%), Q1a (1,32%) и J1 (0,85%). Гаплогруппы с суммарной частотой менее 0.5% на диаграмме не приведены.Данные проценты гаплогрупп среди европейцев учитывают численность коренного населения европейских стран с населением свыше 1 млн. человек и среднюю частоту гаплогрупп по ним из нижеприведенной таблицы, суммарная погрешность не превышает 5%.
Данные гаплогруппы с разной частотой от 0-95% встречаются среди немцев, испанцев, итальянцев, греков, поляков, венгров, французов, албанцев, англичан, ирландцев, шотландцев, португальцев, швейцарцев, чехов, словаков, сербов, хорватов, румынов, белорусов, украинцев, датчан, шведов, норвежцев, финнов, русских, эстонцев, латышей, литовцев и др.
Генетика европейцев
Сами гаплогруппы не несут генетической информации, т.к. генетическая информация находиться в аутосомах- первых 22 пар хромосом. Посмотреть распространение генетических компонент в Европе можно здесь. Гаплогруппы являются всего лишь метками давно минувших дней, на заре формирования современных народов.
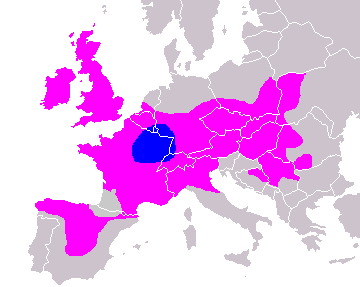
Карта распространения гаплогрупп в Европе из диссертации О. Балановского(только для представленных более 35%)
juliakey.livejournal.com
ГАПЛОГРУППА ИИСУСА ХРИСТА J1 - Дагот Ур
Гаплогруппы чечено-ингушей: J2,J1, F, L3
Попытка создания ДНК-генеалогии Иосифа, Девы Марии и Иисуса Христа
Согласно Священным книгам, примерно 4000 лет назад в Месопотамии родился общий предок будущих евреев и арабов, и по Библии (Ветхий Завет), Торе и Корану имя его было Авраам (Ибрагим). Строго говоря, неизвестно, к какой гаплогруппе Авраам принадлежал, но с конца 1990-х годов популяционные генетики-евреи фактически постулировали, что он относился к гаплогруппе J1.
В принципе, как ранее показал автор настоящего исследования, общие предки евреев и арабов гаплогруппы J2 и R1a тоже жили между 4000 и 5000 лет назад (Клёсов А.А. Славяне, кавказцы, евреи с точки зрения ДНК-генеалогии. М., Книжный мир, 2015, 351 стр.), но какую бы гаплогруппу из трех (J1, J2 или R1a) ни взять, это не изменит выводы настоящего исследования. Поэтому будем считать, что это была гаплогруппа J1, а остальные варианты держать «про запас».
Итак, без малого 4000 лет назад один из обладателей гаплогруппы J1, житель древнего города Ур, одного из древнейших городов Шумера, вышел со своим семейством из города и отправился на северо-запад, в Ханаан, на средиземноморское побережье. Звали его Аврам, и он был потомок Эвера, праправнука Сима, первого сына Ноя, как повествует Библия. С ним были отец Фарра, жена Сарай и племянник Лот. Причины, по которым он оставил родной город, неизвестны. Но известно, что в те времена Ур пришел в экономический упадок, и многие жители, которые не были заняты сельскохозяйственным трудом и не были привязаны к своим земельным владениям, мигрировали из Ура.
Теперь напомним, почему Авраама предпочтительно относят к гаплогруппе J1. На самом деле, поначалу его относили к гаплогруппе J, без детализации субклада. Но потом оказалось, что именно гаплогруппа J1 наиболее представлена у коэнов, они же коханим (что на иврите означает «священники»), причем наиболее представлена в субкладе J1a2b-P58, а именно в количестве 99 человек из 215 тестированных коэнов (46% от всех) (Hammer M.F., Behar D.M., Karafet T.M., Mendez F.L., Hallmark B., Erez T., Zhivotovsky L.A., Rosset S., Skorecki K. (2009) Extended Y chromosome haplotypes resolve multiple and unique lineages of the Jewish priesthood. Hum. Genet. 126, 707-717). Еще 63 коэна показали при тестировании гаплогруппу J2 с субкладами, 29%), из которых почти половина, 31 человек, приходятся на субклад J2a-M410. Гаплогруппу R1a имели 5 коэнов, то есть 2.3%.
Понятно, что коэны, потомки Аарона, в свою очередь, прямого потомка Авраама, если следовать библейской истории, могут относиться только к одному субкладу, поскольку каждый субклад из отмеченных имеет возраст много тысяч лет. Один древний субклад и только один может быть «субкладом Авраама» и далее «субкладом 12 колен израилевых», то есть потомков Авраама, его сына Исаака и его внука (сына Исаака) Иакова. Исходя из максимальной численности коэнов, почти половины, в гаплогруппе J1-P58, будем в дальнейшем считать, что вероятность «гаплогруппы Авраама», а значит, и 12 колен израилевых (в том числе колена Иуды, его потомка царя Давида, и его потомка Иосифа Плотника, о которых речь пойдет ниже) максимальна для этой гаплогруппы.
Иоанн Креститель – сын Елизаветы и Захарии, священника из рода Ааронова (гаплогруппа J1), и родили они его в преклонном возрасте.
Анатолий А. Клёсов,доктор химических наук, профессор
Гаплогруппа J2 зародилась примерно 35000 лет назад на Ближнем Востоке(Шумер, Аккад, Элам, Вавилон, Ассирия) Самая высокая частота мире гаплогруппы J2 находится среди ингушей (88% мужского населения) и чеченцев (56%) мужчин на Северо-Восточном Кавказе.
Об этих поколениях прородителей человечества рассказывают не только Ветхий Завет, Псалтырь, Евангелие
Три J2 найдено на месте захоронения меровингов (романо-франкский переходный период)
Мерови́нги (потомки Иисуса Христа) (фр. Mérovingiens, нем. Merowinger или Merovinger) — первая династия франкских королей в истории Франции. Короли этой династии правили с конца V до середины VIII века на территории современных Франции и Бельгии.
Интересно что вайнахи - чеченцы и ингуши, единственный народ у которого данная древняя гаплогруппа доминирует
Преобладающие носители J2 сицилийцы, ингуши, шугнанцы, чеченцы, ягнобцы, двалы, киприоты, критцы, македонцы, парсы, греки Северного Причерноморья (урумы, румели, эллины).Немало древних средиземноморских и ближневосточных цивилизаций процветали на территориях, где гаплогруппа J2 была преобладающей. Это хатты, хурриты, хетты, этруски, критяне, греки, финикийцы (и их карфагенское ответвление), израильтяне, и в меньшей степени, также римляне, ассирийцы и персы. За ределами Кавказа, самой высокой частоты гаплогруппа J2 наблюдается на Кипре (37%), Крит (34%), Северный Ирак (28%), Сицилия (26,5%)Римляне наверняка способствовали распространению гаплогруппы J2 в пределах своих границ, судя по распределению J2 в Европе (где частота более чем 5%), что имеет любопытное сходство с границами Римской империи. Максимальная концентрация гаплогруппы J2a в мире на острове Крит (32% населения). Субклад J2a3d (M319), предположительно, родом из Крита.Гаплогруппа J2a также достигает высоких частот в Анатолии, на Южном Кавказе, в Чечне и Ингушетии.
Клан Ротшильдов принадлежит именно к этой гаплогруппе Rothschild DNA Y- J2
Интерес к этим цепочкам снипов вызван в первую очередь тем, что субклад J1-P58 ранее приписывался коэнам (коханим), прямым потомкам первосвященника Аарона, брата Моисея, выведшего евреев из Египта примерно 3600 лет назад. Сначала, в конце 1990-х, коэнов просто определяли в гаплогруппу J1 (правда, коэны, носители J2, были не согласны, и считали, и продолжают считать, что у Аарона была гаплогруппа J2). Затем, в ходе наиболее детальной работы 2000-х годов, Hammer et al (2009) выяснили, что по результатам тестирования 215 коэнов они расходятся по 21 субкладу многих гаплогрупп, из которых:
Любомирские (польск. Lubomirscy, укр. Любомирські) — польский княжеский род герба «Шренява», восходящий к началу XVI века. Lubomirski Family is J2b2 +L283
По какому-то уже устоявшемуся стереотипу многие считают, что гаплогруппы J1 и J2 – это еврейские гаплогруппы. Это, конечно, не так, как уже видно из перечня некоторых кавказских этносов выше, они вовсе не евреи. Общие предки гаплогруппы J2 у многих групп на Кавказе уходят на глубину времен 6-8 тысяч лет назад, и они пришли частью с так называемыми урукскими миграциями из Месопотамии, частью из Передней Азии, частью из Средиземноморья. Евреев как таковых тогда вообще не было, евреи и арабы как этно-социальные и религиозные образования появились только 4000-3600 лет назад.
ИНГУШСКИЙ ГЕН L3, L1CПредполагается, что гаплогруппа возникла около 105—85 тыс. лет тому назад.Носители: пакистанцы: калаши, пуштуны, буруши, ингуши, чеченцыПри этом гаплогруппа есть в генофонде Кавказа у чеченцев с частотой до 14%, у ингушей — около 3%Есть версия о происхождении нынешних L3 от фараонов, которые в свою очередь спустились с небес для создания расы людейветвь Л3 е
Ингушский ген FГаплогруппа F — Y-хромосомная гаплогруппа человека. От этой гаплогруппы происходит большинство других гаплогрупп, в общей сложности охватывающих порядка 90 % современного мужского населения планеты.
J2 Эта гаплогруппа действительно очень интересная. Во первых она входит в кластер J-I который является гаплогруппой первых современных людей Европы пришедших на смену неандертальцам. Братская I единственная возникшая в Европе из всех гаплогрупп.Локализуются ли гг территориально среди вайнахов ? Скажем J1 на определенной территории повышенно, L3 также или всё перемешено и территориально никак не связывается ?
Вот в том то и дело что чаще всего перемешано. Например J2, J1, L3 распространены от Ингушетии до Дагестана, если говорить о территории нахов. Но внутри групп есть некоторые нюансы, касающееся веток. К примеру галгаевская ветка в основном локализуется только в галгаевском обществе в районе треугольника Таргим, Хамхи и Эгикал и рядом расположенных аулах. Шатойская ветка локализуется в Шатойском районе--Хал Келой, Саттой, Нуохой и рядом находящихся аулах. Повторяю это не гаплогруппы локализуются, а ветки внутри гаплогруппы. Это скорее всего тейпы, которые имеют одного предка, от которых произошли эти ветки. Также есстественно в каком то районе преимущественно есть одна группа. Например среди Ч1анти пока мы обнаружили только J1, но это большое общество и охвачено мало людей. Например L3 больше расположена в Галанчожском районе и Веденском.
Речь Иисуса на КрестеЭлои, Элои! ламма савахфани?) Одно из речений Иисуса на кресте, цитата из Пс 21:2: "Боже Мой, Боже Мой! для чего Ты оставил Меня?" (Мф 27:46; Мк 15:34). По-евр. эта фраза звучит как: "Эли, Эли, лама азавтани".
Ингуш.язык ломма: послезавтра Чечен.язык послезавтра(lama): ламаИнгуш.язык эьли: князь Ингуш.язык витани: оставил Ингуш.язык аз: я, азах: меня Ингуш.язык лоам: гораФиникийскиц язык ламед: л, гора
akievgalgei.livejournal.com
Образы древних римлян с Волги в артефактах
8-10 апреля 2010 года я ездил по делам в Санкт-Петербург. Посчитал своим долгом сходить в Эрмитаж, захватил с собой фотоаппарат и купил билет на фотосъемку.
В Эрмитаже собрана одна из лучших коллекций в мире оригинальных образцов античного искусства, включая артефакты из Древней Греции, Рима и Боспорского царства. Не мудрствуя лукаво, я стал обходить залы с римскими, греческими и боспорскими экспонатами, тем более что они находятся в одной части Зимнего дворца и располагаются одни за другими. Я фотографировал все подряд, за исключением нескольких всемирно известных статуй богов и бюстов императоров.
Ниже приводится мой фотографический отчет об экскурсии в прошлое.
Я уже несколько лет провожу исследования в области истории и реконструирую события без привязки на политические, религиозные и общепринятые каноны и стереотипы. В залах Эрмитажа я получил полный набор доказательств, подтверждающих мою реконструкцию истории, суть которой заключается в существовании единого вектора развития и расселения человеческой цивилизации из региона Поволжья по всему миру.
Факты упрямая вещь и они говорят о том, что в реальности никогда не существовало массы людей прекрасной, арийской наружности ни в Древней Греции, ни в Риме. Среди миллионов неказистых древних гоминидов встречались отдельные личности, которых можно считать красивыми людьми европейской наружности. Эти малочисленные люди выглядели как арии, остальные были серой массой прототипов современных людей.
Причем все красивые арийцы, как мужчины, так и женщины представляли собой либо богов, либо род царей и их родственников. Нам известно, что их было за тысячи лет всего несколько сот человек. При этом западная цивилизация внушила нам, что все римляне и эллины были подобны богам, лица их были прекрасны, а фигуры атлетического сложения. Ренессанс в Европе в XVI-XVII веках создал многочисленные образцы имитаций древнего искусства. В них запечатлены классические образы древних людей, которых лепили или писали с натурщиков, живших среди авторов произведений. Поздние художники и скульпторы XVIII-XX веков также приложили немалые усилия, изображая в своих произведения «древних» эллинов и римлян, чтобы в свою очередь приукрасить творения Ренессанса. В результате непрерывной работе масонских мастеров за 500 лет мы сегодня думаем, что древние люди были богоподобны, а европейская культура и цивилизация вышла из районов Греции и Италии. Якобы все лучшие образцы искусства были сделаны западноевропейцами в противовес грязному, немытому, страшному и дикому Востоку, включая территорию современной России.
На самом деле это наглая ложь и мистификация. Современная цивилизация распространилась с территории России из регионов Поволжья, Кавказа и Алтая – все предки древних греков и римлян были оттуда родом. Древний Рим располагался в районе селения Селиторное на Волге–Ахтубе. Население древнего города называло его на булгарском языке Идель, на латинском языке Рим, а на египетском и греческом языках Мемфис. Река Тибр есть латинское название реки Ахтуба, от слова туба (труба, канал). Гербом Иделя и Рима была волчица (смотрите статью "Капитолийская волчица из Поволжья"). Там жили семь древних племен, ведущих своё начало от семи праматерей, и город лежал на семи холмах. Городское население было монгольско-булгарской наружности. Мегаполис располагался на пересечении караванных и морских путей запада и востока, севера и юга. Из Поволжья люди пришли в Средиземноморский регион, основав цивилизацию греческих городов-колоний. Древние греки также были монгольско-булгарской наружности. Только элита греков и римлян выглядела, как современные европейцы. Элиту возглавлял род царей Руси, который на булгарском языке назывался Дуло. Со временем элита ассимилировало население, которое стало приобретать черты арийцев.
В II-I веках до нашей эры и в первых веках нашей эры римляне с Волги–Ахтубы осуществили экспансию на запад, опираясь на помощь родственных племен этрусков в Италии и греческих городов-колоний на побережье морей. В IV веке нашей эры Рим был захвачен египетской династией фараонов Нового царства, основанного римскими императорами Гордианами, родом из Африки. В 324 году Рим пал под ударами армий фараона Тутмоса III, потомка Гордианов. В 378-379 годах из Рима ушла первая волна переселенцев, названная в европейской историографии первым гуннским нашествием. Беженцев возглавил Флавий Феодосий, ставшим императором Нового Рима Феодосием Великим. Булгарское имя императора было Арбат, и он являлся царем династии Руси-Дуло или Хонской династии. Именно хоны или гунны были древними римлянами. При переселении они захватили с собой образцы своей культуры, произведения искусства и драгоценности. Эти предметы были перенесены с Волги-Ахтубы в Европу, Балканы и Италию.
Вторая волна переселенцев древнего Рима двинулась в Европу в V веке в 440-450 годах. На этот раз беженцев возглавил внук Флавия Феодосия по имени Флавий Аэций, получивший титул императора с именем Феодосий II. Флавий Аэций это никто иной, как легендарный Аттила, очередной царь из рода Руси-Дуло Хонской династии. Второе «гуннское» нашествие также перенесло массу артефактов и предметов искусства из Древнего Рима в Европу. Римляне (якобы гунны) вновь опустошили Европу, проявив чудеса жестокости и варварства. После Флавия Аэция, называемого «последним римлянином», или Аттилы по булгарски, больше никто в истории не осуществлял подобных грандиозных переселений народов.
Таким незамысловатым образом предметы искусства Рима попали в Европу, в том числе на территорию северной и южной Италии. В центре Италии была Этрурия – государство этрусков, которые имели свое самобытное искусство, похожее на древнеримское с Волги–Ахтубы. Только в XIII-XV веках был осуществлен грандиозный проект католической церкви и западного масонства по созданию имитации Древнего Рима в центре Италии на месте старинного поселения этрусков. Этот Рим существует до сих пор, являясь столицей Италии.
История древнего города – столицы мира – Иделя, Рима, Мемфиса, Мицраима на этом не закончилась. В VII веке древние булгары они же израильтяне окончательно изгнали фараонов с Волги–Ахтубы, и к концу века Мемфис вновь обрел свое старинное имя – Итиль. Со временем Итиль стал столицей могущественного Хазарского каганата. В 965-969 годах русский Великий Князь Святослав с помощью войска варягов и римлян со стенобитными машинами и специальным вооружением захватил Итиль. Хазарский каганат был уничтожен. В XIII веке Итиль получил новую жизнь и название – Сарай-Бату, ставший одной из столиц Золотой Орды. Наконец в XVI веке царь Иван Грозный окончательно уничтожил Сарай-Бату, разобрав его сооружения и стены. Из полученного строительного материала была возведена Астрахань. Во время уничтожения Сарай-Бату длина стен азиатской части города составляла около 18 км при ширине сектора 8 км, а на острове Ахтуба длина стен была около 12 км и ширина сектора 6 км. Фактически древний город Сарай-Бату по размерам не уступал современной Москве. Долгие тысячелетия в Иделе-Риме-Мемфисе-Мицраиме-Итиле-Сарай-Бату жили от 600 000 до миллиона человек. Руины города являются грандиозной кладовой истории, культуры и религии. Масонские ученые старательно избегают проводить там масштабные раскопки. В тех местах процветает только черные кладоискатели.
Доколе возможно мировой элите самозванцев скрывать истину от людей?
20-22.04.2010.
1. Фрагмент римского саркофага с головами на углах I-II век
Обратите внимание на строгость композиции и изысканные позы фигур. При этом посмотрите на мужчин, изображенных по углам саркофага – это монгольско-булгарские лица.
2. Римский саркофаг II век
Также красивая композиция в классическом стиле, однако, мы присмотримся к лицам римлянок на саркофаге и обилию коней в Риме.
3. Крышка римского саркофага I век
Строгая римская композиция на стенке саркофага. Посмотрите внимательно на круглые монгольско-булгарские лица ангелов, а также их крупные тела без талии и шеи.
4. Крышка римского саркофага II век
Другая изысканная стенка римского саркофага. Посмотрите на монгольские лица и тела ангелов и женщины.
5. Фрагмент римского саркофага I-II век
Крупный план одной стенки римского саркофага. Здесь женские лица похожи на европеек, но при внимательном рассмотрении видно, что это тюркские кавказские женщины.
6. Женская голова с крышки саркофага
Крупный план римского женского лица – оно типично булгарское.
7. Вторая женская голова с крышки саркофага
Другой крупный план римской булгарской женщины.
8. Женские головы с крышки саркофага
Вот еще две римлянки с Поволжья.
9. Фрагмент римского алтаря I век
Монгольское лицо медузы-горгоны с римского алтаря.
10. Стенка саркофага, Рим, I век
Монголо-булгарский ангел во всей своей римской красе – круглое лицо, крупное тело без талии, короткие ноги.
11. Портрет римлянки, первая половина I века
Римлянка с тюркско-славянскими чертами лица.
12. Римлянка, II век
Другая римлянка с типично монгольской и кавказской наружностью.
13. Римлянка, начало II века
Еще одна римлянка с Кавказа.
14. Римлянин, II век
Уважаемый римский аксакал с гор.
15. Римлянин, II век, вид сбоку
Профиль аксакала – наденьте на него папаху и отрастите бороду – этому римлянину впору объявлять джихад.
16. Римский ребенок
Милый римский мальчик с монгольско-булгарской внешностью.
17. Римский мальчик
Еще один римский мальчик – короткие ноги и тело, круглое неказистое лицо.
18. Римский сатир
Римский сатир с телом атлета и монгольским лицом
19. Одна из римских богинь
Римская богиня, несмотря на арийское происхождение, удивительно похожа на крупную булгарскую женщину.
20. Бюст Афродиты
Бюст римской булгарской богини. Шея по ширине круглого лица.
21. Голова богини
Голова другой римской булгарской богини – типичной восточной женщины.
22. Богиня Рома, вид сбоку
Скульптура римской богини Ромы. При всем к ней уважении она выглядит как булгарка – широкий таз, короткие ноги и круглое лицо.
23. Богиня Рома, вид спереди
Богиня Рома с другой позиции.
24. Богиня Рома, амазонка 195 год до н.э.
Лицо булгарской женщины – богини Ромы с длинным, большим носом. На обратной стороне монеты она скачет на лошади, как амазонка с Дона.
25. Богиня Рома на слонах, 122 год до н.э.
Римская богиня Рома в упряжке на слонах в «Древнем Риме». Лицо ее также трудно назвать классическим.
26. Богиня Рома или Марс на оленях, 75 год до н.э.
Римская богиня Рома в упряжке из северных оленей. Типичная картина для Италии?
27. Римская богиня
Одна из римских богинь – монгольско-булгарская женщина
28. Статуя греческой богини, первые века до н.э.
Греческий вариант богини – более европеизированная булгарская женщина с коротким, толстым туловищем и короткими ногами.
29. Гробница царей Боспора, Кубань, Крым
Удивительный саркофаг из Боспорского царства на Кубани. Качество скульптуры гораздо выше многих «римских» оригиналов. Композиция идентична известной погребальной плите этрусков.
30. Картер, Италия, IV век до н.э.
Картер римлян – вглядитесь внимательно в круглые монгольские лица на ручках сосуда.
Расположение легендарного древнего города Идель-Рим-Мемфис-Мицраим-Итиль-Сарай-Бату на Волге-Ахтубе.
Пришли результаты ДНК-тестирования. Как и ожидалось мои предки принадлежат к гаплогруппе J2
Другие высокие частота гаплогруппы J2 можно найти во многих других кавказских популяциях, в том числе азербайджанских (30%), грузин (27%), кумыков (25%) и армян (22%). Тем не менее, это очень маловероятно, что гаплогруппы J2 возникла на Кавказе из-за очень низкого генетического разнообразия в регионе. Большинство кавказских народов принадлежат к тому же субкладу J2a4b (снип М67). Высокая частота в данных местах наблюдается в результате эффекта основателя, например распространение вождей и королевских линий через длинную традицию многоженства, практики, что веками русские пытались подавить, после их завоевания Кавказа в 19 веке.За пределами Кавказа, самой высокой частоты гаплогруппа J2 наблюдается на Кипре (37%), Крит (34%), Северный Ирак (28%), Сицилия (26,5%), Ливан (26%), Турции (24%, с пиками 30% в регионе Мраморного моря и в Центральной Анатолии), Южной Италии (23,5%), Болгарии (20%), Албании (19,5%) и материковой Греции (19% без учета северной Греции), а также среди евреев ( от 19 до 25%).Одна четвертая часть у изолированных общин романских языков на Балканах принадлежат к гаплогруппе J2, которая, в сочетании с тем, что они говорят на языке, произошедшем от латинского, предполагает, что они могут иметь большую часть римских линий на Балканах.
История и субклады гаплогруппы J2
Два основных древнейших субклада гаплогруппы J2: J2a (M410, L152, L212/PF4988, L559/PF4986) и J2b (M12, M102, M221, M314).
Ближне-Восточные и европейские J2a
Гаплогруппа J2a значительно присутствует в Италии и является свидетельством в отношении миграции этрусков с Ближнего Востока в страны Центральной и Северной Италии, а также греческой колонизации южной Италии.Финикийцы, евреи, греки и римляне все это способствовало присутствию гаплогруппы J2a в Иберии. Особенно сильная частота J2a и других ближневосточных гаплогрупп (J1, E1b1b, T) на юге Пиренейского полуострова, предполагают, что финикийцы и карфагеняне играли более решающую роль, чем другие народы. Это имеет смысл, учитывая, что они были первыми, основавшими наибольшее количество городов (в том числе Гадир / Кадис, старейший город Иберии), и их поселения почти точно соответствуют более высокой частоте зоны Южной Андалусии.Римляне, конечно, способствовали распространению гаплогруппы J2 в пределах своих границ, судя по распределению J2 в Европе (частота более 5%), которая имеет странное сходство с границами Римской империи.Максимальные концентрации в мире J2a находятся на Крите (32% населения). Субклад J2a4d (M319), вероятно, родом из Крита.
Индийские J2a
В Индии гаплогруппа J2a более распространена среди высших каст. Это можно объяснить ассимиляцией местных (бактрийских) J2a (и возможно G2a) выходцев из Центральной Азии представителями гаплогруппы R1a- индоевропейских воинов, которые мигрировали с Южного Урала (синташтинского культура) и зарекомендовали себя в течение нескольких веков на юге Центральной Азии, непосредственно к северу от Гиндукуша (в том числе Окса цивилизации), прежде чем перейти на завоевание индийского субконтинента. J2a достигли бы южной части Центральной Азии с расширением народов Ближне-Восточной эпохи неолита и смешались с местными охотниками-собирателями принадлежащих в основном к R2 (и, возможно, некоторыми до-индоевропейскими R1a1).
Гаплогруппа J2b
Гаплогруппа J2b имеет совершенно другое распределение в отличие от J2a. Гаплогруппа J2b, вероятно, связана с энеолитом культур Юго-Восточной Европы, и особенно часто встречается на Балканах, в Центральной Европе и Италии, которые составляют примерно масштабы европейской культуры медного века. Её максимальная частота достигается вокруг Албании, Косово, Черногории и Северо-Западной Греции. Гаплогруппа J2b также находится в понтийских степях Северного Кавказа, Центральной Азии и в Южной Азии, особенно в Индии. В очень низкой частоте на Ближнем Востоке, хотя предполагают, что, в отличие от J2a, они не были распространены прогрессивным и непрерывнм распространением образа жизни неолита. По этой причине, как правило, они среди высших каст Индии, считается, что некоторые J2b линии, возможно, были частью индо-арийских вторжений в Южную Азию (3500 лет назад) наряду с гаплогруппой R1a1. Вполне возможно, что меньшинство J2b, G2a3b1 и R1b1a2 из Кавказского региона мигрировали в Волго-Уральском регионе в эпоху ранней бронзы, распространяясь с ними прото-индо-европейский язык и бронзовые технологии в Каспийский степи до расширения этой новой культуры в страны Центральной и Южной Азии.
sunja-edu.livejournal.com
Гаплогруппа G и G2a у осетин, грузин и адыгов
Тип - Y-ДНК Предполагаемая дата появления - 20 000 лет назад.Предполагаемое место появления - Ближний Восток или Юго-Западная Азия Предок - гаплогруппа F Сестринские группы - H, IJ (предок I и J) и K Потомки - G1, G2 и субкладыХарактеристика мутаций - M201 (G), M285 (G1), P287 (G2) с общими субкладами: P16 (G2a1), M406 (G2a3a), G2a3b1 (P303), M377 (G2c)
Типичные представители
(>25%): грузины, адыги, абхазы, абазины, балкарцы, карачаевцы, осетины(дигорцы);(>5%): жители остров Сардиния, Ивиса, Крит, Кипр, Сицилия; австрийцы, шведы, богемцы, венгры, армяне, персы, пуштуны, калаши, турки, южные украинцы, южные немцы, северные итальянцы, обитатели некоторых районов Испании и Франции;Гаплогруппа G — "западнокавказская" Y-хромосомная гаплогруппа человека, определяющаяся маркёром М201. Предположительно возникла около 20 тысяч лет назад на Ближнем Востоке, по другой гипотезе, к югу или к западу от Гималайских гор, в районе Кашмира.
Происхождение гаплогруппы G
Первоначальное её расселение связывается со временем неолитической революции на территории «Плодородного полумесяца», Закавказья и Средиземноморья.Хотя большинство найденных в захоронениях Сибири и Средней Азии скифских скелетов содержат гаплогруппу R1a1, конкретно в местах обитания алан (восточных сарматов) распространена гаплогруппа G, в связи с чем многими современными исследователями гаплогруппа G связывается именно с аланами. Косвенным аргументом в пользу этого служит сохранение гаплогруппы G у осетин, адыгов и у части населения юго-восточной Украины.
Распространение гаплогруппы G
Кавказ
В настоящее время из довольно крупных народов, гаплогруппа G с наибольшей частотой встречается среди осетин (обнаружена у 60 % осетинских мужчин), в Дигорском и Алагирском районах Северной Осетии — до 75% субклад G2a1.Из небольших популяций, очень высока частота встречаемости гаплогруппы G среди шапсугов и казахского рода маджар — около 80 %.Следующими после маджар, шапсугов и осетин по частоте встречаемости данной гаплогруппы идут абхазы и терские казаки — у них она встречается с частотой более 50%В Карачаево-Черкесии, Кабардино-Балкарии, Сванетии и Шида Картли(Грузия) — около 30 %. Известным представителем гаплогруппы G2a1 был Иосиф СталинНа Кавказе к востоку от Осетии — среди чеченцев, ингушей и народов Дагестана гаплогруппа G встречается значительно реже — с частотой 0-5 %. Исключение составляют лезгины и аварцы, у них с частотой около 15% и 10% соответственно.
Славяне
Как сказано выше, гаплогруппа G обнаружена примерно у половины терских казаков. В целом среди русских частота встречаемости гаплогруппы не более 2%, у украинцев не более 3%. Также, с небольшой частотой в 5% встречается среди чехов и 1% у хорватов.Помимо них, в Восточной Европе гаплогруппа G наиболее распространена на юге России и юго-восточной Украине (ок. 5%) — что некоторыми исследователями считается результатом включения традиционно обитавших там алан в состав пришедшего позднее восточнославянского населения.В целом среди русских и украинцев частота встречаемости гаплогруппы G — 3-4 %.
Гаплогруппа G в Европе
В других местах Европы гаплогруппа G довольно часто встречается в материковой Греции, северной Испании и Италии, на Крите, Сардинии и в Тироле (до 15%). На юге Германии и в Венгрии встречается с частотой до 7%.В остальной Европе гаплогруппа G встречается редко (менее 5% в целом по континенту) и представлена в центральной её части отдельной подгруппой G2c.
Азия
В странах Азии гаплогруппа G с частотой 10-20 % встречается у иранских персов, пуштунов (афганцев) и калашей. Также встречается у турок и армян с частотой 11 %, у иракских арабов с частотой около 10 % и у курдов. Менее распространена в Пакистане, Индии (брахманы). Обнаружена у курейшитов. Самое большое скопление разновидностей гаплогруппы находится в Ливане (Финикия).
Субклады гаплогруппы G
Гаплогруппа G рапространена на обширной территории от Европы до Азии еще в доисторические времена и имеет много субкладов. Два основных субклада это G2a1 и G2a3b.
Гаплогруппа G определяется мутацией в M201. Первый человек,с мутацией M201 , как полагают, появился около 30.000 лет назад (1200 ~ поколений), вероятно в Закавказье, но, возможно, также вдоль восточной границы на Ближнем Востоке, возможно, и на Памире. У него было относительно мало потомков.
Распределение гаплогруппы G
Гаплогруппа G, вероятно, образовалась вблизи Кавказских гор, где она имеет наибольший процент современной концентрации и разнообразия. Ее члены рассеялись в Центральной Азии, Европе и на Ближнем Востоке во время наступления неолита. Те, кто пошел на восток имеют очень небольшое количество потомков по мужской линии в Китае, Индонезии, на Тайване, Филиппинах, и полинезийских островах. Те, кто отправился на север у мужчин-линии потомков в Сирии (арабы), Россия (Aдыги), Узбекистан (татары и каракалпаки), Монголия и Западный Китай (уйгуры). Те, кто пошел на запад и север представлены сегодня в Европе. Гаплогруппа G является наиболее распространенной в Кавказском регионе, в том числе около 60 процентов осетин и 30 процентов грузин, кабардинцев и балкарцев, и уменьшение процента в Азербайджане (18 %) и Армении (11 %).Гаплогруппа G представляет собой одну из значительных ветвей неолитического населения древнего Ближнего Востока. Гаплогруппа G хорошо представлена там и сегодня. У израильских евреев (9,8 процента), Турция (9,2 процента), Египет (9 процентов), Палестина ( 8,9 процента), Ливана (6 процентов), Иордании (5,5 процента), Сирия (4,8 процента), и Саудовская Аравия (4,5 процента). Это она была, вероятно, одной из основателей популяции древних евреев. Сегодня, около 10% евреев, как ашкеназов так и сефардов, принадлежат к гаплогруппе G.Только от 1 до 3 процентов у современных европейцев встречается гаплогруппа G, с градиентом от юго-востока (наиболее распространенная) и на северо-запад и север (наименьшее общее количество). Есть концентрация на Сардинии (14 процентов), о. Ибица (13 процентов), северо-центральной части Италии (10 процентов), Корсика (11,8 процента), на Крите (10,9 процента), северо-востоке Испании (8,3 процента), Мальта (8 процентов), Австрийский Тироль (7 процентов), Португалии (7,3 процента), и Чешская Республика (5,1 процента). Средиземноморские концентрации могут указывать поселения финикийцев и колонистов карфагенской империи. Гаплогруппа G, вероятно, распространялась римлянами, как вербовка солдат и движения купцов. Её современное распространение в Европе появляется отслеживать тесно с границами Римской империи.
Гаплогруппа G2
Гаплогруппа G2 определяется мутацией в P15. Похоже, возникла на Кавказе или в Закавказье, но дата является неопределенной. Сегодня, G2 наиболее часто встречается в Европе и на Ближнем Востоке. Одна карта распространения этой гаплогруппы показывает концентрацию в центральной Италии, распространяя на север, в швейцарских Альпах. Эта группа возможно, связана с потомками этрусков. Гпплогруппа G2 имеет две подгруппы, G2a и G2b, определяемые мутациями в Р16 и M286, соответственно. G2a имеет подгруппу G2a1, что имеет дополнительные мутации на P17 и P18.
Гаплогруппа G2a3b1
Гаплогруппа G2a3b1 определяется мутацией P303. В эту группу входят большинство европейских мужчин в гаплогруппе G. Основатель, появился около 5 тысяч лет назад, вероятно, где-то на Ближнем Востоке, возможно, в Турции или Иране. Его основные подгруппы, как полагают, разделились , возможно, 4 тысячи лет назад, и распространились на Европу между 1500 и 2500 лет назад. На протяжении многих лет было распространено мнение, что эта мутация маркера для сарматских солдат, проходящих службу в римских легионах возле в Великобритании. Вдальнейшем было обнаружено также что значительная концентрация данного субклада на континенте, и это вероятно связано с аланским наследием, варварскими племенами, что попали в Европу в 5-ом столетии, сначала, как римские солдаты, а затем и в качестве союзников вестготов. Однако, в течение последних нескольких лет, тестирование сарматских потомков и Алан на родине вблизи Кавказа показал, что нет близких матчей европейских мужчин в той же группе. Т.е. если аланы были носителями гаплогруппы G, то у низ мог быть только субклад G2a1.Теперь ясно, факты не подтверждают Аланского или Сарматского происхождения. Вместо этого, этот субклад, возможно, была распространен в Европе купцами, как вариант - еврейскими. Теория остается спорной. Существует еще одна подгруппа P303, которая включает в себя большое число евреев-ашкенази, и этот субклад имеет свою крупнейшую европейскую концентрацию на острове Ибица, который, как известно, показывает значительное население крипто-евреев. Однако, нет доказательств того, что группа в целом была когда-нибудь что-нибудь более конкретным, чем Ближний туземцев Восточной.
Гаплогруппа G2a3b1b1
Гаплогруппа G2a3b1b1 определяется мутациями в L42/S146 и L43/S147. Эти мутации были обнаружены в генетических образцах из Джастин Хауэри-Сванстром, испытанных в 23andMe. FamilyTreeDNA впоследствии разработала коммерческий тест. Около 30 процентов DYS388 = 13 у мужчин, которые проверены на маркер L42 и показали положительные результаты. Они содержат самые различные значения маркера, указав, что это очень старый SNP. В настоящее время считается, что 25 процентов мужчин G2 из Швейцарии, вероятно, члены этой группы.Эти мутации, кажется, почти столь же древний, как и P303, может быть, датируемые 4500 лет назад. Взятые вместе, они, вероятно, включают маркер для древних популяций Этрурии и Реции.
Этруски и гаплогруппа G2a
Городская культура этрусков предшествовали росту Рима. Этруски были из числа не-индо-европейских народов. Их культура развивалась в северной и центральной Италии где то около 800 года до н.э. Этрусская цивилизация процветала в Тоскане и долине реки Пo до прихода галлов и роста Римской республики. Римляне ассимилировали этрусков около 500 до н.э.Происхождение этрусков является спорным. В пятом веке до н.э. греческий историк Геродот считал, что предки этрусков возникли с колонией лидийцев из Анатолии (Турция). (Геродот, история (ок. 430 до н.э.), 1.94). Другие древние историки придерживались других мнений. По Дионисию Галикарнасскому (1 век до н.э.), этруски были коренными в Италии.
Распределение Гаплогруппа G2a
Претензия Геродота на лидийское происхождение была спорной с самого начала, но поддерживается и в настоящее время на основе проведенных генетических исследований. Ряд исследований показали более высокий уровень ДНК гаплогруппы G в Тоскане в поддержку теории Анатолийского происхождения. Исследования митохондриальной ДНК у современных тосканцев и древних этрусков также указывают происхождение на Ближнем Востоке. Археология и изучение ДНК тосканских пород крупного рогатого скота предполагает что этруски прибыли в Италию около 1200 до н.э. В карте выше, мы видим, значительные концентрации G2a в Кавказском регионе. Также обратите внимание на концентрации в древней Анатолии (современная Турция), в центральной Италии, и в древней северной Италии. На карте ниже, мы видим всю широту распространения ареала гаплогруппы G.
Реция
Были племена, к северу от Альп, которые были завоеваны римлянами. Они жили на территории нынешней Восточной и Центральной Швейцарии (это район Верхнего Рейна и Боденского озера), на юге Баварии и Верхней Швабии, Форарльберг, большая часть Тироля, и часть Ломбардии. Сегодня в этом регионе обнаружены относительно высокие частоты гаплогруппы G2 по сравнению с остальной Европой.Жители Реции считают, что их предки были этруски, которые были изгнаны из равнин реки По в Италии при вторжении галлов (386 г. до н.э.).К тому времени, Реция и её жители появляется впервые в истории, они были полностью объединены с кельтскими племенами и поселились в том же районе. В начале шестого века Реция была оккупирована готами, и в 9-м веке она была включена в состав франкского государственного устройства. Даже если гаплогруппа G преобладала среди ранних жителей Реции, учитывая эту историю, это не удивительно, что другие гаплогруппы сейчас преобладают в том же районе.
Вышеизложенная информация была частично взята из книги Спенсера Уэллса, путешествие Генетическая Одиссея (Random House, 2004).Проект гаплогруппы G Подробное филогенетическое дерево Y гаплогруппдДНК из ископаемых останков различных эпох
haplogroup.narod.ru
Забытая реальность — Миф о древних ДНК-генеалогических линиях африканцев (часть 1)
В академической литературе за последние четверть века регулярно появляются утверждения о том, что африканские ДНК-линии «самые древние», хотя основания для того не приводятся. И понятно, почему не приводятся – их просто нет. Давайте для начала разберемся, что такое «древние ДНК-линии», понимая под этим цепочки снипов Y-хромосомы. Но разбираться особенно и не надо, потому что у всех мужчин на Земле, африканцев и неафриканцев, во всяком случае, у всех, кто делал тест на ДНК, снип-цепочки Y-хромосомы по древности одинаковы. У всех эти цепочки снипов тянутся от общих предков с шимпанзе, орангутангом, гориллой, макакой и другими древними общими с нами предками. Ниже – характерный пример, процитированный еще в 2012 году в журнале Advances in Anthropology (Klyosov, Rozhanskii, Ryabchenko): Сравнение фрагмента нуклеотидной последовательности Y-хромосомы
человека, шимпанзе, гориллы, орангутанга и макаки Видно, что в 90% случаев нуклеотидные последовательности показанных фагментов Y-хромосомы совпадают. А у шимпанзе и человека совпадают 93 нуклеотида из 97, то есть в 96%. Так что у всех нас ДНК-корни древние, и африканцы в этом отношении никак не выделяются. Хорошо, а насколько древние? Антропологи оценивают время расхождения линии будущего человека от линии будущего шимпанзе в 4-6 миллионов лет назад, это же повторяют и генетики, хотя поиски статей с конкретными расчетами результатов не дали. А вот результаты расчета с помощью автоматического калькулятора Килина-Клёсова (ККК, 2015), иллюстрация увеличивается по клику: Общий предок человека и шимпанзе жил, если округлить расчеты, 4.2±1.5 миллионов лет назад. 16-маркерный гаплотип шимпанзе был извлечен из геномной базы данных, как описано в той статье в Advances in Anthropology (2012), а в качестве обобщенного гаплотипа человека брался экстраполированный предковый гаплотип по всем неафриканским гаплогруппам, пересчитанный на времена 100 тысяч лет назад. Впрочем, можно было и не пересчитывать, вот – время до общего предка современных носителей гаплогруппы А00 и шимпанзе, иллюстрация увеличивается по клику: Оно равно 4.4±1.4 миллионов лет назад, что практически одинаково в пределах погрешности расчетов. Те же времена дают все основные гаплогруппы серии А, например: • А0а-V150 имеет общего предка с шимпанзе 4.6±1.7 млн лет назад; • A1b1-M32 (Чад и Турция) 4.3±1.4 млн лет назад; • A1b1-M32, M13 (Британия) 4.2±1.4 млн лет назад, а также • гаплогруппа В, 4.4±1.4 млн лет назад. Короче, в отношении «древних ДНК-линий» африканцы ничем от неафриканцев не отличаются. И те, и другие тянут свои ДНК-линии от общего предка с шимпанзе. И у тех, и у других цепочки снипов уходят на глубины в сотни тысяч и миллионы лет. К этому мы еще вернемся. Может, под «древними африканскими ДНК-линиями» попгенетики и прочие генетики подразумевают древние времена жизни их общих предков, и полагают, что они намного более древние, чем у неафриканцев? Придется их огорчить, ситуация противоположная. Африканские предки группы А (популяций, субкладов) почти все совсем недавние. Возьмем, например, гаплогруппу А00, которая среди гаплогрупп считается самой древней. Да, для нее цепочка снипов насчитывает – по данным, приводимым YFull – 1684 снипов, что при 144 годах на снип (это число приводит YFull) дает 242500 лет от начала гаплогруппы. Действительно, YFull приводят близкий «возраст» А00, 235 тысяч лет. Но откуда эта цепочка снипов ведет – никто не знает. Может, из Африки, может, с Кавказа, может, из Сибири, может, из Европы – неизвестно. Почему-то считается, что именно из Африки. Но это опять «по понятиям», правда, откуда появившимся, непонятно, извините за игру слов. Древнейшие ДНК находили в Европе, на Русской равнине, в Сибири, а в Африке ни разу, правда, и исследований таких не было. Так откуда эти «понятия» появились? А так, стиль попгенетиков. Кстати, о возрасте гаплогруппы А00. Калькулятор КК дает для базовых гаплотипов А0 и А00 возраст общего предка (с округлением) 217000±87000 лет. Это – возраст А00 или немного старше. Так что датировка по снипам воспроизводится по гаплотипам в пределах погрешности расчетов (иллюстрация ниже увеличивается по клику). Так вот, об общих предках африканских популяций, субкладов группы А. Они почти все по времени очень мелкие, часто всего несколько сотен лет. Иногда – между 1000 и 2000 лет. Самые древние – примерно 5000-7000 лет. И опять – где эти предки жили, никто не знает. Взглянем на гаплогруппу А00. В литературе (Mendez et al., 2013) описаны 11 гаплотипов в 95-маркерном формате (без 16 мультимаркерных гаплотипов) носителей гаплогруппы А00, все из племени Mbo из Камеруна, и еще один – афроамериканец. Расчетов в статье, конечно, нет. Все 11 гаплотипов содержат 29 мутаций, что дает 29/11/0.161 = 16 условных поколений, но с учетом того, что 45 аллелей (из 1045 аллелей) авторы определить не смогли, то до общего предка на самом деле 17 условных поколений, то есть 425±90 лет. Итак, сама гаплогруппа А00 образовалась более 200 тысяч лет назад, причем неизвестно где, а общий предок носителей А00 из Камеруна жил примерно 425 лет назад. Если добавить к серии из Камеруна еще гаплотип афроамериканца той же гаплогруппы, который отличается от предкового гаплотипа камерунцев на 11 мутаций, то есть на 11/0.161 = 68 → 73 условных поколения, то есть на 1825 лет, то их общий предок, камерунцев и афроамериканца, жил примерно (425+1825)/2 = 1125 лет назад. Примеров недавних общих предков разных мужских популяций гаплогрупп класса А, которые сейчас живут в Африке, можно привести немало. В большинстве этих популяций гаплотипы или одинаковы или почти одинаковы, даже для протяженных гаплотипов. С доступными гаплотипами гаплогруппы А00 мы уже разобрались, им четыре сотни лет. Взглянем теперь на Проект FTDNA гаплогруппы А: • В гаплогруппе/субкладе А0-V150 имеются два 25-маркерных гаплотипа, оба идентичны друг другу, то есть их общий предок жил не более 300-400 лет назад. Других гаплотипов в 25-маркерном формате или выше нет. • В гаплогруппе A0a-L981 (L994, V148, V166) между двумя доступными 67-маркерными гаплотипами есть 8 мутаций, что помещает их общего предка на 900 лет назад. • В гаплогруппе A0b-V166, L896 шесть 37-маркерных гаплотипов имеют общего предка 475±160 лет назад. • В гаплогруппе A1a-M31 («европейский кластер») пять 37-маркерных гаплотипов имеют общего предка 1175±290 лет назад. • В гаплогруппе A1a-M31 (все неизвестного происхождения) пять 37-маркерных гаплотипов имеют общего предка 1880±390 лет назад. • В гаплогруппе A1a-M31 (из 18 гаплотипов – 14 неизвестного происхождения, два из Англии, один из Швейцарии, один Кейп Верде) все 37-маркерные, все имеют общего предка 730±130 лет назад. • В гаплогруппе A1a-M31 («британский кластер») 19 гаплотипов в 67-маркерном формате имеют общего предка 740±120 лет назад. В итоге, 50 гаплотипов в 37-маркерном формате четырех предшествующих серий (А0а-М31) имеют общего предка 9270±970 лет назад. Но надо обратить внимание, что это четыре серии гаплотипов, разбросанные по разным континентам. Это – не одна «ДНК-линия», это – четыре разных ДНК-линии, с четырьмя общими предками, каждый «глубиной» от 700 до 1900 лет. Ну и где, позволительно спросить, «древнейшие ДНК-линии в Африке»? Это – миф, ничем не обоснованный. Ископаемых ДНК нет, древних линий, как мы видим, нет, к тому же, в Африке вовсе не гаплотипы группы А явяются доминирующими. Доминирует гаплогруппа E1b; в Камеруне и Чаде, и отчасти в Нигерии, доминирует гаплогруппа R1b, поэтому неудивительно, что в Африке наблюдается «высокое генетическое разнообразие», только это не от древности, а от обилия носителей разных гаплогрупп, прибывших в Африку в разные времена. Да и вообще, как можно серьезно обращать внимание на «генетическое разнобразие» в открытых системах, какой является Африка? Как я уже не раз отмечал, самым «генетически разнообразным» городом мира является, наверное, Нью-Йорк, но вряд ли кто будет серьезно утверждать, что Нью-Йорк – древнейший город мира. Кроме, наверное, популяционных генетиков, с их примитивными и бездумными критериями. Откуда же появился этот миф о «древности африканских линий»? Датировок ведь их до последнего времени не было, да и датировки бывают, как мы убедились, нескольких видов – цепочек мутаций, уходящих далеко вглубь, до приматов; цепочек мутаций, ведущих ко временам образования соответствующих гаплогрупп; и датировок общих предков современных носителей соответствующих гаплогрупп и субкладов. Ни одна из этих датировок не «укоренена» территориально, то есть мы не знаем, на какой территории, на каком континенте образовалась та или иная гаплогруппа. Мы можем только набирать сведения и складывать фрагменты общей картины, но Африка в этом отношении находится на одном из последних мест по накопленным знаниям. Но фантазий, натяжек и откровенных фальсификаций про них со стороны попгенетиков – хоть отбавляй. Собственно, это характерно для популяционных генетиков, но они уроки из принципиальных ошибок не извлекают. Еще не так давно они утверждали, что гаплогруппа R1b образовалась в Европе 30 тысяч лет назад, и сначала приняли в штыки, а потом напрочь игнорировали сведения и датировки ДНК-генеалогии, что R1b прибыла из Южной Сибири длинным миграционным путем в Европу примерно 4800 лет назад (Klyosov, 2009; Klyosov, 2012). Называли это «псевдонаукой», особенно в этом усердствовали попгенетики Балановские. Разворот произошел в 2013 году, когда неподалеку от Байкала нашли ископаемую гаплогруппу R с археологической датировкой 24 тысячи лет назад. Лед тронулся, и попгенетики, не моргнув глазом, не повинившись и не цитируя статьи с датировками ДНК-генеалогии, стали повторять то, что уже было опубликовано нами несколько лет назад. Например, в недавней статье Batini, Jobling и других (2015) датировки гаплогруппы R1b в Европе уже даются как R1b-L11 4510 лет (доверительный интервал 3920-5160 лет назад) и R1b-P312 4210 лет (доверительный интервал 3700-4790 лет назад). YFull дают датировки R1b-U106 4900 лет (доверительный интервал 4400-5400 лет назад), R1b-P312 4600 лет (для TMRCA, доверительный интервал 4300-5000 лет назад). К сожалению, приходится повторять, что попгенетики уроков из своих ошибок не извлекают. Ситуация повторяется с Африкой. Данных нет, или данные говорят о другом, но их придумывают и искажают. Зачем это им нужно? Для нормального человека звучит невероятно, но это ментальность вассалов, которые лебезят перед сюзереном. Сюзерен решает, кого пригреть, кого пригласить на конференцию, кому дать грант, кого взять в коллективную статью, ввести в комиссию или комитет, в общем, в инфраструктуру. Без сюзерена вассал – никто, и вассал это прекрасно понимает. А зачем это нужно сюзерену? О, там большая политика. Вассала к этим вопросам не допускают. Вассал знает, что если он публично усомнится в африканском происхождении человечества, то его дни сочтены. Поэтому он, вассал, готов разорвать на части любого, кто сомневается или выставляет данные против. А поскольку он разорвать на части не может, то цель – найти компромат, состряпать ложь, передернуть, исказить – других способов или аргументов у вассала нет, во всяком случае, научных. Посмотрим на схему гаплогрупп и субкладов группы А, приведенную согласно классификации ISOGG, в сокращенном виде: В этой схеме – источник принципиальной путаницы попенетиков. Они не смогли увидеть и осознать, что эта лесенка субкладов представляет собой серию вилок, которые периодически расходятся на «африканские» (по сегодняшнему месту жительства) и неафриканские ветви. Опишем кратко эти вилки, а потом рассмотрим их подробнее – когда они образовались, и кто являются потомками субкладов по обе стороны тех вилок. Итак, вилок всего четыре. Первой от ствола, уходящего вглубь эволюционного дерева человечества (точнее, его Y-хромосомы), отходит вилка, знаменующая, видимо, начало Homo sapiens – расхождение на А00 и А0-Т. Вторая – расхождение А0-Т на А0 и А1. Третья – расхождение А1 на А1a и A1b. Четвертая – расхождение A1b на A1b1 и ВТ. Если упрощенно – то каждая вилка расходится на африканскую (по нынешнему месту жительства) и неафриканскую стороны. А00, А0, A1a и A1b1 уходят в Африку, противоположные стороны вилки A0-T, A1, A1b и BT остаются вне Африки. «Упрощенно» – потому что вилка на А00 и A0-T образовалась еще до расхождения на африканцев и неафриканцев (это расхождение произошло примерно 160 тысяч лет назад), а впоследствии многие неафриканцы перебирались в Африку – это и многочисленные носители гаплогрупп E1b, J2, R1b, и так далее. Среди них – миллионы современных жителей Западной Африки (берберы, жители Камеруна и Чада), египтяне, потомки буров в Южной Африке, и так далее. Немало жителей Европы и Ближнего Востока имеют гаплогруппу A1a, как недавно показывал И.Л. Рожанский в статье на Переформате. Да и вообще носителей группы А в Африке относительно мало – часто на уровне единиц процента или того меньше. Рассмотрим вилки подробнее. Первая вилка – расхождение предшественника Homo sapiens (или Homo sapiens в начальной стадии, примерно 230 тысяч лет назад) на гаплогруппы А00 и А0-Т. Нисходящих субкладов от А00 пока не обнаружено, и ее нынешние носители обнаружены только в Африке в количестве 11 человек (см. выше), с общим предком несколько сотен лет назад, а также несколько афроамериканцев с одинаковыми гаплотипами. Снипы гаплогруппы А0-Т, напротив, имеются у всех неафриканцев, в том числе у всех нас, читателей Переформата, хотя только у мужчин. И, разумеется, у всех африканцев, носителей гаплогрупп E1b, J2, R1b, В и других, которые прибыли в Африку относительно недавно. Но помимо них снипы группы А0-Т должны быть и у всех африканцев, носителей групп А, потому эти снипы предшествуют разделению африканских и неафриканских линий (примерно 160 тысяч лет назад). Впрочем, речь идет о промежуточных снипах в цепочках наших и африканских снипов, носителей А0-Т как терминальных снипов пока ни у кого не выявлено, ни в Африке, ни вне Африки. Поэтому называть его «африканским» никакого смысла не имеет. К тому, что у всех нас, мужчин, есть множество снипов гаплогруппы А0-Т – ниже приведен список части из 587 снипов этой гаплогруппы, из которых у меня есть L1090, L1121, L1124, L1125, L1127, L1235 и многие другие: Мой список снипов BigY CTS10042, CTS10627, CTS10834, CTS10847, CTS10976, CTS10993, CTS11148, CTS11184, CTS11411, CTS11468, CTS11530, CTS1169, CTS11720, CTS11734, CTS11853, CTS11913, CTS11948, CTS12010, CTS1211, CTS1277, CTS1278, CTS12947, CTS12972, CTS1340, CTS1367, CTS1415, CTS1416, CTS1417, CTS1419, CTS1422, CTS1619, CTS1738, CTS1795, CTS1907, CTS2134, CTS262, CTS2626, CTS2636, CTS2637, CTS2638, CTS2639, CTS2663, CTS 2680, CTS2891, CTS2988, CTS2992, CTS3067, CTS3075, CTS3229, CTS3230, CTS3231, CTS3316, CTS3321, CTS3358, CTS3402, CTS3441, CTS3527, CTS3534, CTS3548, CTS3551, CTS358, CTS3607, CTS3622, CTS3649, CTS3650, CTS3654, CTS3697, CTS3775, CTS3848, CTS3943, CTS3984, CTS4259, CTS4465, CTS4862, CTS501, CTS5287, CTS5423, CTS5454, CTS5508, CTS5577, CTS5648, CTS570, CTS5936, CTS5979, CTS623, CTS6918, CTS6985, CTS7191, CTS7275, CTS7278, CTS7400, CTS7500, CTS7650, CTS7659, CTS7941, CTS8008, CTS8127, CTS8350, CTS8507, CTS8508, CTS8591, CTS8626, CTS8665, CTS8797, CTS8816, CTS8896, CTS9018, CTS903, CTS9108, CTS9510, CTS9515, CTS9539, CTS9596, CTS9733, CTS9739, CTS9754, CTS9779, F1050, F1209, F1224, F132, F1586, F184, F185, F2957, F3032, F313, F3159, F3166, F3185, F3194, F3197, F3337, F3561, F3564, F3644, F3650, F3692, F4099, F686, F719, F773, F82, F937, F989, IMS-JST002612, IMS-ST022448_3, IMS-ST022448_6, IMS-ST022449_3, IMS-ST022449_6, IMS-ST022451_2, K511, L1002, L1004, L1005, L1009, L1013, L1028, L104, L105, L1053, L1060, L1061, L1062, L1067, L1071, L108, L1089, L1090, L1095, L1098, L1099, L110, L1101, L1105, L1112, L1116, L1118, L112, L1120, L1121, L1123, L1124, L1125, L1127, L1129, L113, L1132, L1135, L1136, L1137, L114, L1142, L1143, L1145, L1150, L1155, L1179, L1185, L1120, L1225, L1235, L132, L1346, L1347, L1352, L1480, L1492, L15, L16, L168, L248, L268, L313, L350, L352, L413, L418, L438, L440, L449, L457, L468, L469, L498, L543, L58, L604, L721, L74, L741, L747, L76, L760, L768, L777, L779, L781, L819, L82, L821, L851, L875, L882, L962, L969, L970, L977, L986, L989, M168, M173, M198, M207, M213, M235, M251, M294, M299, M306, M417, M42, M420, M448, M449, M45, M459, M511, M512, M513, M514, M515, M516, M526, M74, M89, M9, M94, P108, P128, P131, P132, P133, P134, P135, P136, P138, P139, P14, P140, P141, P143, P145, P146, P148, P149, P151, P157, P158, P159, P160, P161, P163, P166, P187, P19_1, P207, P224, P225, P226, P228, P229, P230, P231, P232, P233, P234, P235, P236, P237, P238, P239, P242, P243, P244, P245, P27.1_2, P280, P281, P282, P283, P284, P285, P286, P294, P305, P316, PAGES00026, PAGES00081, PAGES00083, PF1345, PF1378, PF1389, PF1396, PF1402, PF1403, PF1405, PF1407, PF1408, PF1411, PF1412, PF1414, PF1415, PF1418, PF1419, PF2589, PF2590, PF2591, PF2608, PF2610, PF2611, PF2614, PF2615, PF2616, PF2619, PF2620, PF2621, PF2622, PF2626, PF2629, PF2631, PF2634, PF2635, PF2637, PF2640, PF2642, PF2647, PF2651, PF2653, PF2658, PF2660, PF2683, PF2685, PF288, PF2690, PF2694, PF2695, PF2700, PF2709, PF2716, PF2718, PF2722, PF2723, PF2729, PF2731, PF2732, PF2736, PF2737, PF2739, PF2742, PF2747, PF2748, PF2749, PF2752, PF2753, PF2754, PF2755, PF2756, PF2757, PF2758, PF2770, PF2772, PF2988, PF349, PF3494, PF3495, PF3496, PF3497, PF3500, PF4522, PF5459, PF5469, PF5477, PF5483, PF5488, PF5490, PF5498, PF5500, PF5503, PF5513, PF5861, PF5862, PF5863, PF5864, PF5869, PF5870, PF5871, PF5872, PF5876, PF5881, PF5882, PF5885, PF5886, PF5888, PF5889, PF5895, PF5908, PF5911, PF5914, PF5916, PF5919, PF5920, PF5936, PF5940, PF5943, PF5944, PF5945, PF5949, PF5951, PF5953, PF5954, PF5955, PF5956, PF5957, PF5958, PF5964, PF5965, PF5967, PF5970, PF5971, PF5975, PF5978, PF5980, PF5981, PF5982, PF5984, PF5985, PF5991, PF6013, PF6016, PF6047, PF6054, PF6056, PF6057, PF6066, PF6069, PF6071, PF6072, PF6073, PF6114, PF6116, PF6118, PF6121, PF6132, PF6133, PF6143, PF6153, PF6156, PF6158, PF6159, PF6160, PF6162, PF6163, PF6165, PF6168, PF6169, PF6170, PF6172, PF6179, PF6181, PF6194, PF6195, PF6201, PF6204, PF6211, PF6212, PF6213, PF6214, PF6215, PF6216, PF6217, PF6218, PF6220, PF6222, PF6230, PF6231, PF6233, PF6512, PF682, PF7514, PF7522, PF7527, PF7530, PF7531, PF7532, PF7534, PF7540, PF7542, PK1, PR1220, PR202, PR2119, PR5761, PR855, rs161492_1, V221, V238, V241, V250, Y2902, Y32, Y33, Y34, Y36, Y783.2, YSC0000057, YSC0000077, YSC0000081, YSC0000107, YSC0000108, YSC0000109, YSC0000110, YSC0000193, YSC0000350, YSC0000913, Z1331, Z280, Z282, Z5997, Z6030, Z6032. Фрагмент из списка снипов гаплогруппы A0-T (приведены 182 снипа из 587) Понятие «вилка» не стоит понимать буквально, А00 и А0-Т могли образоваться с разницей в десятки тысяч лет. По данным YFull, до начала А00 насчитывается 1684 снипа, до начала А0-Т – 587 снипов, но возраст обеих гаплогрупп почему-то выставлен как одинаковый, 235 тысяч лет назад. На такую повторяющуюся «дихотомию» в презентации данных YFull мы уже не раз обращали внимание. В любом случае, искать у нас, читателей Переформата, снипы гаплогруппы А00 бесполезно, нет их у нас, ни одного из тех 1684 снипов. Но важно то, что А0-Т образует очередную вилку, на гаплогруппы А0 и А1. Вторая вилка – расхождение А0-Т на А0 и А1. Потомки А0 сейчас живут в Африке, потомки А1 – мы с вами, читатели Переформата. У всех нас в цепочках снипов есть снипы субклада А1 (см. ниже), но это опять не значит, что наши предки вышли из Африки. Это опять промежуточные снипы, и никого с терминальными снипами А1 пока не выявлено, ни в Африке, ни вне Африки. Поэтому называть его «африканским» никакого смысла тоже не имеет. По данным, приведенным YFull, к A0 ведет 899 снипов (это 129460 лет, если снип образуется в среднем раз в 144 года, как считали те же YFull), к А1 – 62 снипа (8900 лет), возраст А0 не выставлен, но для А1 выставлено 146300 лет «возраста». Опять загадка, как это может быть. Судя по этим данным, на протяжении 100 тысяч лет между А00 и А0-Т, гаплогрупп между ними вовсе не образовывалось, и к чему те 62 снипа относятся – непонятно. Но здесь важнее то, что А0 – это «африканская» гаплогруппа (опять по нынешнему месту жительства), а А1 – неафриканская. На схеме выше показано семь субкладов гаплогрупы А0, и снипов никаких из них у нас, читателей Переформата, нет, а снипы гаплогруппы А1 есть у нас всех (я не буду больше повторять, что это относится к обладателям Y-хромосомы, то есть к мужчинам, и что «читатель Переформата» относится к неафриканцам, потомкам гаплогруппы ВТ, которые не перебрались на жительство в Африку). Повторю, что «африканских» снипов А0 у нас нет, можно не искать, нет ни одного из 899 тех снипов. Не выходили наши предки из Африки, или, во всяком случае, не являются потомками ни А00, ни А0. Надо сказать, что в Африке носителей гаплогруппы А0 обнаружено немного. Scozzari et al (2012) указывали, что только после долгих поисков нашли трех (!) носителей гаплогруппы А0, все в Камеруне, и отмечали, что эта гаплогруппа имеет в Африке очень низкую частоту встречаемости. Ниже приведен список из 62 снипов гаплогруппы А1, из которых у меня есть L986, L989, L1002, L1009, L1112, V238, V241, V250. Остальных нет потому, что не все снипы у каждого определяются. Список снипов гаплогруппы A1 Третья вилка – расхождение А1 на A1a и A1b. Потомки A1a сейчас живут в Африке и по всему миру (в частности, в ряде европейских стран и на Ближнем Востоке). Снипы А1b есть у всех нас, читатели Переформата. Но это опять не значит, что наши предки вышли из Африки. Это опять промежуточные снипы, и никого с терминальными снипами А1b пока не выявлено, ни в Африке, ни вне Африки. Поэтому называть его «африканским» опять никакого смысла не имеет. По данным, приведенным YFull, к A1a ведет 495 снипов (это 71 тысяча лет, если снип образуется в среднем раз в 144 года), к А1b – 44 снипа (6300 лет), но почему-то им опять было выставлено по 132600 лет «возраста». Опять загадка, на каком основании так делается. Хотя носители гаплогруппы A1a разбросаны по всему миру (поэтому в любом случае это не «африканская» гаплогруппа, даже по такому условному критерию, как современное место жительства), их мало. И.Р. Рожанский уже приводил в своей статье «Африканцы на севере Европы. Африканцы ли?», что гаплогруппа A1a-M31 чрезвычайно редкая, и даже в Африке ее наличие исчисляется в лучшем случае 4.4% (в Западной Африке – Гвинея-Бисау, Гамбия, Сенегал, Мали, Нигер, Марокко), и 0.4% среди афроамериканцев. Этому же вторит статья Scozzari et al. (2012), что им удалось найти в Африке для геномных иследований только двоих (!) носителей A1a-M31. Ниже приведен список из 44 снипов гаплогруппы А1b, но в той системе, которую использует YFull. В классификации ISOGG в гаплогруппе A1b насчитывается 47 снипов, и ни один не пересекается по индексам с приведенными ниже. Из этих 47 снипов у меня есть L989, L1009, L1013, L1053, P108, V221. Остальных нет, потому что, как указывалось выше, не все снипы у каждого определяются. Список снипов гаплогруппы A1b (согласно классификации, используемой YFull) И вот здесь снипы гаплогруппы A1b приоткрывают загадку. Воспроизведем еще раз фрагмент лесенки снипов гаплогруппы A1: Мы видим, что от нее вилкой отходят субклады A1a и A1b. Мы уже узнали, что А1а встречается в Европе, на Ближнем Востоке, в Африке, причем нет никаких данных, что он изначально был африканским. Субклад A1b встречается у всех неафриканцев как промежуточный, в цепочке снипов. Субкладов, нижестоящих от A1b, а это A1b1, A1b1a, A1b1a1, A1b1a1a, A1b1a1a1, A1b1a1a2, A1b1a1a2a, A1b1a1a2b – у неафриканцев нет, возможно, потому, что параллельно с A1b1 образовался субклад BT (по даным YFull примерно 126 тысяч лет назад), и A1b1 ушли в Африку, а ВТ остался вне Африки. То есть у неафриканцев имется основная линия A1b > BT, и потомки ее составляют все гаплогруппы от В до Т. И вдруг выясняется, что нижестоящая линия, в субкладе A1b1a1- M23, P3, обнаружилась в Южной Африке среди носителей койсанских языков. Но это – довольно далекий по цепочке вниз субклад, который, по-видимому, отошел от неафриканцев в древние времена. Иначе говоря, не неафриканцы отошли от африканцев, а наоборот, африканцы отошли от неафриканцев, мигрировав в Африку. Опять – не выходили наши предки из Африки. Было наоборот: современные африканцы – потомки неафриканцев. Четвертая вилка – расхождение A1b на A1b1 и BT. A1b1 и нисходящие субклады считаются африканскими – по нынешнему месту проживания его носителей, хотя родительская гаплогруппа A1b (как промежуточный субклад в цепочке снипов) насчитывает не менее 2-3 миллиардов неафриканцев, а нижеследующие субклады от A1b1 и ниже – обнаружены всего у нескольких африканцев. Вот и считайте, кто от кого произошел. Ответ ясен – африканцы от неафриканцев. Повторю – не выходили наши предки из Африки. Итак, потомки A1b1 сейчас живут в минимальном количестве в Африке, а потомки гаплогруппы ВТ – это все неафриканские гаплогруппы от В до Т. По данным YFull, к A1b1 ведут всего 26 снипов, к субкладу A1b1a ведут всего 2 снипа, к A1b1a1 – 11 снипов, ни одного из них у нас нет. Датировок ни к одному из нисходящих от A1b1 субкладов в списке YFull нет, и понятно, почему – их носители крайне редко встречаются, да и данных нет. Снипы ВТ есть у всех нас, читатели Переформата. Список снипов гаплогруппы A1b1 (согласно классификации, используемой YFull) К сводной гаплогруппе ВТ ведут 403 снипа, и многие из них есть у нас. У гаплогрупп категории А их, естественно, нет. Датировка ВТ по данным YFull – примерно 126 тысяч лет, хотя 403 снипа – это примерно 58 тысяч лет. Некоторые могут подумать, что эти снипы – от родительской гаплогруппы A1b, но та датирована той же компанией YFull примерно 133 тысячами лет назад, то есть всего на 7000 лет раньше. Опять остается неясным, к чему те 403 снипа относятся, и откуда тянутся. Самая загадка в том, как «параллельные» гаплогруппы A1b1 и BT, от одного общего предка A1b, и к которым идут 26 и 403 снипа, соответственно, могут оказаться с одинаковой датировкой – 126 тысяч лет. Ниже приведен частичный список снипов сводной гаплогруппы ВТ. Как уже отмечалось, многие из них есть у каждого неафриканского мужчины. У меня есть, например, L418, L604, L962, L970, L977, PF1408 и другие. Фрагмент из списка снипов гаплогруппы BT (приведены 166 снипов из 403) Итак, первая часть этой статьи безоговорочно показывает, что «африканские ДНК-линии» совсем не древние, в противовес тому, чем нас пичкают популяционные генетики уже 30 лет. Если вести «древность» от общих предков с приматами, то африканские линии ничем не отличаются от неафриканских, имеют одну и ту же глубину. Если считать от времени жизни общих предков современных мужских популяций – по племенам или по территориям, то африканские ДНК-линии весьма мелкие, намного мельче, чем неафриканские. Если считать от образования гаплогрупп, то никто не знает, на каких территориях или континентах они образовались, поэтому присваивать им названия «африканские» – некорректно. Более того, носителей этих гаплогрупп, категории А, в Африке очень мало. К этому могут быть разные причины – прошли бутылочные горлышки популяции, вымерли, или прибыли в Африку относительно недавно – большого значения не имеет, они не «древние», в том смысле, который вкладывают попгенетики. В итоге имеем то, что изображено на заглавной иллюстрации поста. Так что с этим разделом закончили – нет в Африке «древних ДНК-линий». Но есть такое понятие, как совокупные ДНК-линии. Это тогда, когда сами ДНК-линии мелкие, но они – ошметки древних раскидистых деревьев гаплогрупп и субкладов. Мы видим только верхушки, но экстраполяция верхушек, в данном случае гаплотипов этих «ошметок», позволяет увидеть, когда жили общие предки этих разбросанных гаплотипов. Мы уже видели это выше – сравнение гаплотипов «мелких» популяций гаплогрупп А00 и А0 показывает, что они расходятся на астрономические расстояния. Сами «ошметки» имеют общих предков всего несколько сотен лет назад, но они расходятся на полмилиона лет, и отсюда следует, что их общие предки жили около 250 тысяч лет назад. В этом отношении африканские ДНК-линии отличаются от неафриканских. Рассмотрим это подробнее, в этом упрятана одна из основных загадок человечества. Загадка была впервые сформулирована в 2012 году, в статье (Klyosov, Rozhanskii, Advances in Anthropology, 2, No. 2, 80-86). Суть ее состояла в том, что все ДНК-линии современных гаплогрупп от В до Т, то есть неафриканских гаплогрупп, сходятся к общему предку, который жил 64000±6000 лет назад, хотя разрозненные линии африканских (по нынешнему месту жительства) гаплогрупп сходятся к общему предку 132 тысяч лет назад (хотя сами разрозненные линии датируются временами жизни общих предков всего лишь 5500, 5000 и 600 лет назад). При этом все неафриканские линии (включая гаплогруппу В) отстоят исключительно далеко от африканских линий, ясно показывая, что неафриканские линии не могли произойти от африканских. Например, базовые гаплотипы гаплогрупп А0 и В отстоят друг от друга на 270 тысяч лет (между ними 18 мутаций в «медленных» 22-маркерных гаплотипах; для сравнения, базовые гаплотипы R1a и R1b отстоят друг от друга всего на 8 мутаций, то есть примерно на 54 тысячи лет, и это помещает их общего предка на 27 тысяч лет назад), иллюстрация увеличивается по клику. Ясно, что если между гаплогруппами А0 и В 270 тысяч лет, то вторая никак не могла образоваться от первой в течение всего периода существования Homo sapiens, не говоря о 70 тысячах лет назад, которые попгенетики отводят для времени образования гаплогруппы В из А. Это аналогично примеру выше для R1a и R1b, в котором 54 тысячи лет расстояния между ними никак не могут допустить, что одна образовалась из другой; они обе образовались от одного общего предка, которым в данном случае являлась гаплогруппа R1. Гаплогруппа А0, далеко отстоящая от гаплогруппы В, здесь не исключение. Подобные огромные расстояния отделяют гаплогруппу В от любых гаплотипов в субкладах категории А, например, 250 тысяч лет от A0a-L981, 280 тысяч лет от A1b-M32, 310 тысяч лет от европейского кластера гаплогруппы A1a-M31, 360 тысяч лет от другого кластера той же гаплогруппы, 320 тысяч лет от арабского кластера той же гаплогруппы, 350 тысяч лет от A0a-V150, 356 тысяч лет от A0a-L979, 346 тысяч лет от A0a1a-V151, и так далее. Самое большое расстояние отделяет гаплогруппу В от А00 – 450 тысяч лет, то есть их общий предок жил примерно 225 тысяч лет назад, на утренней заре Homo sapiens. Такая же картина наблюдается и для гаплотипов всех гаплогрупп от В до Т. Например, растояние от гаплогруппы С до А0-L896 составляет 357 тысяч лет, до A0a1-V150 – 300 тысяч лет, и так далее. Но расстояние от В до С составляет всего 120 тысяч лет, то есть их общий предок жил примерно 60 тысяч лет назад. Это находится в полном соответствии с картиной, что гаплогруппа В образует единый кластер с гаплогруппами от В до Т, и не является по происхождению «африканской» гаплогруппой. Видимо, уже всем ясно, что гаплогруппа В не образовалась из какой-либо гаплогруппы категории А (поскольку нет гаплогруппы А как таковой, есть А00, А0-Т, А0, А1, А1a, A1b и так далее), а все они ведут свое начало от общего предка. Этот общий предок жил примерно 160 тысяч лет назад, как показано в упомянутой выше статье (Klyosov, Rozhanskii, Advances in Anthropology, 2, No. 2, 80-86). Если исходить из датировок, приведенных на сайте YFull, то основатели гаплогрупп А00 и A0-T жили раньше этого времени, причем гаплогруппа А00 осталась в известном смысле тупиковой, нисходящие от нее субклады и прочие ДНК-линии неизвестны. A0-T прошла своими снипами как к африканцам, так и к неафриканцам, как показано выше, поскольку образовалась еще до разделения на африканскую и неафриканскую ветви. Разделение на эти ветви соответствует временам непосредственно до образования гаплогрупп А0 и А1, причем А0 ушла в Африку, А1 осталась вне Африки. Впоследствии носители многих гаплогрупп мигрировали в Африку, например, A1a и нижестоящие субклады, A1b1 и нижестоящие субклады, поэтому их снипов у нас нет, как описано выше. Все это описано в целом в упомянутой выше статье (Klyosov, Rozhanskii, Advances in Anthropology, 2012), которая, кстати, набрала рекордное количество просмотров (около 90 тысяч) и скачиваний (около 20 тысяч), но академическое сообщество предпочло ее не заметить. Слишком многое она меняет в представлениях. Основная иллюстрация в той статье следующая: Номенклатура в этой иллюстрации несколько устаревшая, в ней нет гаплогруппы А00, которая в 2012 году была неизвестной, но суть ее остается правильной. Гаплогруппа альфа – это условная гаплогруппа, от которой пошло разделение эволюционного Y-хромосомного дерева человечества на африканскую (слева) и неафриканскую линии. В настоящее время все гаплогруппы от В до Т, то есть гаплогруппы неафриканского происхождения, сходятся к условной же бета-гаплогруппе, которой на самом деле является сводная гаплогруппа ВТ. Загадка – куда делась связка между альфа- и бета-гаплогруппой, протяженностью в сто тысяч лет? И, кстати, куда делись африканские ДНК-линии, которые на левой части диаграммы полностью экстраполированы, от общих предков всего от тысячи до нескольких тысяч лет давности, вглубь времен? Со времени публикации той статьи прошло уже более трех лет, в базах данных и в Проектах с тех пор накопилось много гаплотипов, в том числе протяженных, 111-маркерных, и появилась возможность проверить количественные выводы статьи. То, что расхождение африканской и неафриканской ДНК-линий произошло примерно 160 тысяч лет назад, мы уже проверили. Перейдем ко времени жизни общего предка всех неафриканских гаплогрупп – от В до Т. Ниже – дерево гаплотипов этих гаплогрупп. Оно построено по 111-маркерным гаплотипам, и сами гаплогруппы показаны по окружности дерева. Для каждой из 18 гаплогрупп использовали 10-25 гаплотипов, взятых произвольно из выборки, всего 403 гаплотипа nd 111-маркерном формате. Гаплогруппы категории А на дереве не оставили, так как они явно «вылезали» из кругового дерева, поскольку, как мы уже показали, они удалены по мутациям от всех других гаплогрупп. Видно, что все гаплогруппы от В до Т образуют достаточно единую систему ДНК-линий (выраженную здесь через 111-маркерные гаплотипы), так что про некую «африканскую гаплогруппу В» говорить не приходится. Она такая же «автохтонная африканская», как R1b – «автохтонная европейская». Это застарелая болезнь попгенетиков – то, что видят сейчас, значит, так было всегда. Болезнь называется «отсутствие научного подхода», или «отсутствие научной школы», что, впрочем, одно и то же, а если по-русски, то «думать не умеют». 403 гаплотипа в 111-маркерном формате, то есть почти 45 тысяч аллелей – это крепкий орешек, если считать вручную. Более того, за десятки тысяч лет в них накапливаются десятки тысяч мутаций, помимо того, огромное количество возвратных мутаций, что делает 111-маркерную систему такой древности неустойчивой, и неприемлемой для расчетов, поскольку датировки значительно занижаются. Поэтому для данных случаев разработана 22-маркерная панель из самых «медленных» маркеров. Они настолько устойчивы во времени, что для всех 403 гаплотипов за десятки тысяч лет маркер DYS472 вообще не мутировал ни разу, DYS436 – всего 30 раз, DYS590 – 42 раза, и так далее. Сравните – маркер DYS390 за это же время должен мутировать 2270 раз, то есть в среднем 5.6 раз на гаплотип. При таких частотах мутации расчет вклада обратных мутаций уже практически невозможен, система нестабильна. А вот «медленная» 22-маркерная панель с малыми скоростями мутаций позволяет проводить расчеты на десятки и сотни тысяч лет вглубь, и даже на миллионы лет, как было показано выше на примере расчета времени жизни общих предков человека и шимпанзе. Более того, разработанный калькулятор Килина-Клёсова проводит расчеты по каждому маркеру отдельно, используя квадратичный метод, и поэтому не требует введения поправок на возвратные мутации. В итоге ДНК-генеалогия получила незаменимый метод расчета массивных серий гаплотипов в любом формате и любой древности общего предка (Килин и Клёсов, Вестник Академии ДНК-генеалогии, 2015, т. 8, № 3, 321-375, http://dna-academy.ru/kilin-klyosov/). Расчет дерева из 403 гаплотипов, приведенного выше, показал, что общий предок гаплогрупп от В до Т жил примерно 63 тысячи лет назад (результат без округления – 63036 лет, что, конечно, с такой точностью совершенно излишне). Это почти точно совпадает с рассчитанной ранее (Klyosov and Rozhanskii, 2012) величиной 64±6 тысяч лет. Теперь осталось подойти к решению сформулированной в той статье загадки – повторим ее опять: куда же делись предки Y-хромосомных линий в Африке ранее 5-10 тысяч лет назад, и у неафриканцев ранее 64±6 тысяч лет назад? Причем у африканских линий группы А можно косвенным путем, а именно при сравнении протяженных гаплотипов с «неглубокими» общими предками, достичь времен жизни общих предков 132 тысячи лет назад, а у неафриканцев, как их гаплотипы ни комбинировать и ни сравнивать, глубже 64±6 тысяч лет уйти не получается. Более древних общих предков как корова языком слизнула. Что же это за корова была такая? Некоторые предположения в цитированной выше статье сделаны были – упомянуто возможное последствие извержения вулкана Тоба в Индонезии, около 70 тысяч лет назад. Не исключено было резкое похолодание, возможно, как результат извержения того же вулкана, причем, по мнению специалистов, ранг катастрофичности глобального похолодания намного выше, чем извержения вулкана, но это все оставалось довольно абстрактными предположениями. И вообще – чтобы извержение вулкана в Индонезии привело к гибели почти всех людей на планете – это кажется маловероятным, но все это «по понятиям». Нужно было мнение специалиста, пусть мнение и спорное, неоднозначное, но главное – подтвержденное конкретными данными. И такой специалист по данной проблеме есть. Знакомьтесь – Валерий Павлович Юрковец, профессиональный геофизик, палеоклиматолог, специалист в области влияния климатических катастроф, в том числе импактного характера, на миграции древнего человека, член Академии ДНК-генеалогии с начала ее образования несколько лет назад. Продолжение часть 2 Анатолий А. Клёсов, доктор химических наук, профессор Источник
xn--80aaacvi7aqjpqei0jvae5b.xn--p1ai
Ближневосточные гаплогруппы J1 J2 E1b1b1 G2a и др. Описание и связь с археологическими культурами
Важная информация
Информация о происхождении и этнической принадлежности гаплогрупп на этом сайте не должна рассматриваться как неопровержимый факт, но, как это часто бывает в науке, в качестве модели в процессе постоянной эволюции на основе современных знаний и понимания (авторов). Всякий раз, когда прогресс генетики не может предоставить неопровержимые ответы, мы попытались предоставить наиболее вероятные и логичные гипотезы на основе археологических, исторических и лингвистических доказательств. Эта страница постоянно обновляется, чтобы идти в ногу с недавними исследованиями и дает дополнительное понимание и исправление возможно ошибочных теорий.
Гаплогруппа G
Гаплогруппа G, как полагают, возникла на Ближнем Востоке в конце палеолита, возможно раньше, уже 30 000 лет назад. В то время все люди были охотниками-собирателями, и в большинстве случаев, жили в небольших кочевых или полукочевых племенах. Члены этой гаплогруппы, возможно, были тесно связаны с развитием сельского хозяйства в раннем Леванте и в той части плодородного полумесяца, начиная с 11500 лет назад до настоящего времени. Древние Y-ДНК G2a обнаружены также в четырех культурах европейского неолита (ЛБК в Германии, Remedello в Италии и кардиальной керамики в Юго-Западной Франции и Испании), и все результаты дали G2a, что является убедительным доказательством, что сельское хозяйство возникло на Ближнем Востоке и было распространено носителями гаплогруппы G.Гаплогруппа G2a составляет от 5 до 10% населения средиземноморской Европы, но довольно редко встречается в Северной Европе. Единственным местом, где гаплогруппы G2 превышает 10% населения в Европе- Кантабрия, Швейцария, Тироль, на юге центральной Италии (Молизе, Центральная и Южная часть Апеннинского п-ва), Сардиния, северная Греция (Фессалия) и Крит - все горные и относительно изолированные регионы.Есть несколько теорий о происхождении гаплогруппы G2a в Европе.
Горные пастухи неолита
Хронологически первая гипотеза о появлении неолитических фермеров и скотоводов из Анатолии в Европе попадает на период между 9000 и 6000 лет назад. В этом сценарии Закавказские мигранты принесли с собой коз и овец, которые были одомашнены к югу от Кавказа около 12000 лет назад. Это объясняет, почему гаплогруппа G является более распространенной в горных районах, будь то в Европе или в Азии.Географическая непрерывность гаплогруппы G2a из Анатолии в Фессалии на итальянский полуостров, Сардинии, южной и центральной Франции и Иберии уже предполагает, что G2a могут быть связаны с культурой Кардиальной керамики (5000-1500 до н.э.). Древние ДНК тесты проводились на скелетах со стоянки ЛБК в Германии, а также стоянки культуры Кардиальной керамики в южной Франции (Лангедок-Руссильон) и подтвердили, что неолитические фермеры в Европе принадлежали в первую очередь к гаплогруппе G2a .В настоящее время гаплогруппа G2a находится в основном в горных районах Европы, например, Кантабрия (более 10%), на севере Испании, Швейцарии (10%), Австрии (8%), Овернь (8%) в центральной Франции, горных районах Чехии (от 5 до 10%) и Уэльсе (4%). Это может быть потому, что закавказские фермеры стремились на холмистую местность, похожую на свою прародину, которая возможно, хорошо подходит для разведения коз. Но наиболее вероятно, что фермеры G2a бежали от захватчиков бронзового века, таких, как индо-европейцы гаплогрупп R1b и R1a и нашли убежище в горах. Предполагаемая родина гаплогруппы R1b1a2 и прото-индо-европейские миграции связываются с Северной Анатолией, и/или Закавказьем. Кавказ сам по себе является пиковой точкой по распространению гаплогруппы G. Поэтому, вполне возможно, что меньшинство кавказских мужчин, принадлежащих к гаплогруппе G (и, возможно, J2) интегрировались в R1b сообщества, пересекли Кавказ и зарекомендовали себя на северных и восточных берегах Черного моря где-то между 7000 и 5000 до н.э. Те прото-индо-европейцы кто принадлежал к R1b1b2a1 и G2a3b1a до завоевания Европы начали на Балканах около 4000 г. до н.э. и распространились по всему Атлантическому побережью от Иберии до Британских островов куда заселились около 2000 г. до нашей эры. Вопреки этому, G2a* и G2a3*, наиболее распространены в горных районах, G2a3b1a находится в низкой частоте по всей Европе, даже в Скандинавии и России. Более того, G2a3b1 также находится в Индии, особенно среди высших каст. Евразия, объединенная наличием G2a3b1 по всей Европе и Индии является очень сильным аргументом в пользу индоевропейского происхождения. Слияние возрастов G2a3b1 также соответствует времени индо-европейской экспансии в эпоху бронзы.Распределение гаплогруппы G в Европе, Северной Африке и на Ближнем Востоке
Расширение сельского хозяйства с Ближнего Востока в Европу (9500-3800 до н.э.)
Период римского расширения и гаплогруппа G
Вполне вероятно, что G2a прибыли в Европу в эпоху неолита или бронзового века и римляне в дальнейшем способствовали распространению субклада по всей Италии, которая относительно богата на G2a. Миграции в пределах Римской империи, вероятно, способствовали умеренному повышению G2a на север в Галлию и Британию, также частота гаплогруппы G наоборот уменьшается с расстоянием от границы Римской империи. Гаплогруппа G крайне редко обнаруживается в Северных и Балтийских странах в настоящее время, несмотря на то, что сельское хозяйство достигло тех регионов, примерно в то же время, как и Великобритании или Ирландии. Это может быть просто совпадением, так как лесные низменности северной Германии, Польши и Северной Европы оказались беднее в металлах и не привлекли бы рабочих бронзового века с Закавказья. Северо-Восточная Европа также имеет довольно скромную частоту R1b, что еще больше усиливает связи между этими двумя гаплогруппами.
Аланский субклад гаплогруппы G2a1
Единственной этнической группой, которая имеет большинство гаплогруппы G2a1 в настоящее время являются осетины на Кавказе, в современном Республики в составе РФ - Северной Осетии-Алании. Считается, что они связаны непосредственно с аланами, племенами из центральной Азии, связанными с древними сарматами. Средневековое королевство Алания было расположено на Северном Кавказе, в современной Грузии и Осетии.G2a наблюдается в несколько более высоких частотах в Пикардии и Фландрии, чем в соседних регионах. Была гипотеза, что G2a была доставлен в северную Францию и Бельгию аланами, которые преодолели путь по всей континентальной Европе во время варварских вторжений в 5 веке и основали ненадолго королевство на севере Франции.Тем не менее, если есть аланские G в Европе, безусловно, они должны принадлежать к другим субкладам, отличным от миграций неолитических и бронзового веков (не G2a3). G2a1 как наиболее распространенный субклад на Кавказе в настоящее время, сравнительно недавно был в составе аланских миграций (с генетической точки зрения). На самом деле, G2a1 были обнаружены на всем протяжении маршрута миграции алан (Венгрия, Франция, Испания), а также в Великобритании (сарматский элемент?), но вряд ли где-нибудь еще.
Скифская гаплогруппа G1
Римляне, как известно, осуществляли набор скифских и сарматских всадников в свои легионы. По словам С. Скотт Литтлтон в своей книге от Скифии до Камелота, несколько рыцарей Круглого Стола были скифского происхождения, а также легенда о Святом Граале сама возникла в древней Скифии. Эта гипотеза также положена в 2004 году в основу фильма о Короле Артуре, который появляется с приходом скифо-римской кавалерии в Великобритании. Тем не менее, скифы были степными людьми и с большей вероятностью принадлежат к гаплогруппе R1a1-Z93. Если скифы принадлежали гаплогруппе G, то они, предположительно, G1, а не G2a. Это могло бы объяснить рассеянные случаи G1 в северо-западной Европе, хотя и G2a2, которые также были обнаружены в Великобритании и Анатолии, являются еще одним потенциальным кандидатом.
Гаплогруппа J (Y-ДНК)
Гаплогруппа J является ближневосточной гаплогруппой, разделенной на условно северную гаплогруппу J2 и южную гаплогруппу J1. Гаплогруппа J2 на сегодняшний день имеет наибольшее распространение и разнообразие в Южной Европе.
Гаплогруппа J2
Гаплогруппа J2 зародилась в Северной Месопотамии и распространилась на запад, в Анатолию и в южную Европу, а также на восток, в Персию и Индию. Гаплогруппа J2 возможно связана с древними этрусками (минойская цивилизация), греками, южной Анатолией, финикийцами, ассирийцами, вавилонянами.В Европе гаплогруппа J2 достигает своей максимальной частоты в Греции (в частности, на острове Крит, Пелопоннес и Фракия), южной и центральной Италии, южной Франции, южной Испании. Древние греки и финикийцы были главной движущей силой распространения J2 вокруг западного и южного Средиземноморья.Гаплогруппа J2, как полагают, прибыла в Грецию из Анатолии между неолитом и бронзовым веками. Распространение гаплогруппы J2 в Европе
Географическое распределение гаплогруппы J2a имеет сильную корреляцию с распространением сельского хозяйства в северной Месопотамии (где её пики), и далее к Анатолии, Греции, всего Ближнего Востока, Ирана, Афганистана, Пакистана и Западной Индии. Значительное присутствие гаплогруппы J2 в Италии связано с ранними миграциями ранее этруссков с Ближнего Востока в центральную и северную Италию, а также с греческой колонизацией юга Италии.Финикийцы, евреи, греки и римляне способствовали распространению гаплогруппы J2a в Иберии. Особенно сильные частоты гаплогруппы J2a и других ближневосточных гаплогрупп (J1, E1b1b1, T) на юге Пиренейского полуострова, указывают, что финикийцы играли более важную роль, чем другие народы. Это имеет смысл, учитывая, что финикийцы / карфагеняне были первыми, кто пришли, основав наибольшее количество городов (в том числе Гадир / Кадис, древний город Иберии), а их поселения почти в точности соответствуют более высоким частотам гаплогруппы J2a.Римляне наверняка способствовали распространению гаплогруппы J2 в пределах своих границ, судя по распределению J2 в Европе (где частота более чем 5%), что имеет любопытное сходство с границами Римской империи.Максимальная концентрация гаплогруппы J2a в мире на острове Крит (32% населения). Субклад J2a3d (M319), предположительно, родом из Крита. Гаплогруппа J2a также достигает высоких частот в Анатолии, на Южном Кавказе, в Чечне и Ингушетии.
Индийские ветви гаплогруппы J2a
В Индии, гаплогруппа J2a чаще встречается среди высших каст и снижается по частоте с понижением кастового уровня. Это может быть объяснено как ассимиляцией местных J2a (и R2), выходцами из Центральной Азии - индоевропейскими воинами R1a1-Z93, которые произошли из Средней Азии (синташтинская культура) и зарекомендовали себя в течение нескольких веков на юге Центральной Азии, к северу от Гиндукуш, прежде чем перейти на завоевание индийского субконтинента. J2a достигли югa Центральной Азии с расширением в эпоху неолита и смешались с местным населением охотников-собирателей, принадлежащих в основном к R2 (и, возможно, древних R1a*).
Гаплогруппа J2b
Мутация образующая субклад J2b вероятно возникла в Греции или в Анатолии, как и у гаплогруппы E-V13 (см. ниже), с которой она тесно связана. Распространение J2b и E-V13 примерно соответствует древней греческой и римской сферы влияния. Помимо Европы, J2b также можно найти по всей Индии, но только в умеренных частотах между Европой и Индией, это означает, что, в отличие от J2a, у неё было не прогрессивное и непрерывное распространение, как и следовало ожидать от распространения сельского хозяйства. По этой причине, а также потому что, как правило, гаплогруппа J2b распространена среди высших каст Индии, считается, что некоторые J2b линии могло быть частью индо-арийского вторжения в Южную Азию (3500 лет назад) вместе с R1a1-Z93. Вполне возможно, что меньшинство J2b, G2a3b1 и R1b1a2 из Кавказского региона мигрировали в Волго-Уральском регионе в начале бронзового века (см. историю R1b), распространяясь с их прото-индо-европейским языком, бронзовыми технологиями и домашними животными до Каспийской степи и расширения этой новой культуры в Центральной и Южной Азии (см. R1a1).
Гаплогруппа J1
Гаплогруппа J1 является ближневосточной гаплогруппой, которая, вероятно, зародилась в Юго-Восточной Анатолии, близ озера Ван в центре Курдистана. Восточная Анатолия является регионом, где козы, овцы и крупный рогатый скот были впервые одомашнены на Ближнем Востоке. Гаплогруппы J1 почти наверняка связана с распространением скотоводства на всей территории Ближнего Востока и Европы. Гаплогруппу J1 можно разделить на две основные группы: субклад J1c3 (P58), а также другие субклады J1 (J1 *, J1A, J1b, J1c1 и J1c2).Гаплогруппа J1c3 (J-P58) на сегодняшний день является наиболее распространенным субкладом J1. Это типично семитский субклад, составляющий большинство населения Аравийского полуострова, где на его долю приходится около 40%-75% мужских линий. J-P58 также является гаплогруппой коэнов. Примерно половина всех коэнов относятся к субкладу J-P58. В еврейской Библии общим предком всех коэнов определяется Аарон- брат Моисея.Гаплогруппа J1c3, как полагают, распространилась с восточной Анатолии в Левант, и с гор Загроса на Аравийском полуострове в конце последнего ледникового периода (12000 лет назад) с сезонной миграцией скотоводов. Возникновение ислама в 7 веке нашей эры играет важную роль в расширении J1c3 из Аравии на Ближнем Востоке, а также в Северной Африке, и в меньшей степени в Сицилии и на юге Испании.Другие субклады гаплогруппы J1 менее хорошо изучены в связи с их гораздо более низкими частотами. Большинство J1 на Кавказе, в Анатолии и в Европе- это не семитская J1c3. Другие типы J1, скорее всего, распространились в Европу в эпоху неолита. Гаплогруппа J1 особенно часто встречается в горных районах Европы (за исключением Альп и Карпат) - Греция, Албания, Италия, центральной Франции, и самые труднодоступные части Иберии (Астурия, Кантабрия, Кастилия-Ла-Манча), а также в Португалии и Андалусии.Как и гаплогруппа G2, J1, возможно, были основными линиями, которые могли привести домашних животных в Европу. J1 достигает максимальной частоты на восточном Кавказе, и в некоторых этнических группах составляет абсолютное большинство (кубачи, кайтаки, даргинцы), в то время как другие имеют чрезвычайно высокий уровень G2 (шапсуги, северные осетины). Большинство этнических групп на Северном Кавказе, имеют от 20 до 40% каждой гаплогруппы, которые на сегодняшний день доминируют на Кавказе. На Южном Кавказе (Грузия, Армения, Азербайджан), гаплогруппа J2a является одной из основных, но также высоки частоты J1 и G2a.Распределение гаплогруппы J1 в Европе, на Ближнем Востоке и в Северной Африке
Гаплогруппа E1b1b1
Гаплогруппа E1b1b1 (ранее E3b) представляет последний крупный поток миграции из Африки в Европу. Считается, что впервые E1b1b1 появились в районе Африканского Рога и Южной Африки примерно 26 000 лет назад и разошлись на Ближнем Востоке в эпоху мезолита. На европейском континенте они имеет самую высокую концентрацию на северо-западе Греции, Албании и Косово, а затем концентрация уменьшается при удалении от Балкан, в остальной Греции и Западной Турции. За пределами Европы, также в большинстве стран Ближнего Востока, Северной и Восточной Африке, особенно в Марокко, Ливия, Египет Йемен, Сомали, Эфиопии и Южной Африке гаплогруппа E1b1b1 имеет очень высокое распространение.
Разве E1b1b1 могли переселиться прямо из Северной Африки в Европу в связи с изменением климата?
До сих пор неясно, когда гаплогруппа E появилась в Европе. Последние тесты ДНК из неолитических стоянок в южной Германии и южной Франции не показывали след E1b1b1. Это свидетельствует о более позднем прибытии, либо к концу неолита / энеолита, либо в эпоху бронзы. В случае отсутствия Y-ДНК из неолита Греции, Южной Италии и Иберии, ничего не исключает возможности того, что гаплогруппа E1b1b1 присутствовала в этих регионах с неолита, мезолита или даже позднего палеолита. В Северной Африке носители E1b1b1 пересекли Средиземное море (вероятно, в нескольких независимых волнах миграции) в любое время с последнего ледникового максимума (около 20000 лет назад), или последнего опустынивания Сахары, который начался, когда муссон отступил на юг 6200 лет назад.В последний ледниковый максимум, уровень моря был на 120 метров ниже, чем сегодня, и Гибралтарский пролив был всего несколько километров, что позволяло использовать даже самые примитивные плоты, чтобы пересечь его легко. Является ли это просто совпадением, что последние следы неандертальцев в Иберии (на самом деле в Гибралтаре) с даты 24000 лет назад, незадолго до последнего ледникового максимума? Может ли их исчезновение являться результатом поглощения Homo Sapiens из Северной Африки? Последние неандертальцы иберийского п-ва показывают некоторые признаки гибридизации с Homo Sapiens. В то время как Homo Sapiens бесспорно колонизировали Европу из стран Ближнего Востока, борьбы с текущим колонизациями с северо-запада Африки, вполне вероятно, тоже были. Это объясняет, почему существует так много Северо-Западно африканских E-M81 в Португалии и северо-западной Испании, которая не подтверждается никакими историческими миграциями, ни любыми археологически очевидными неолитическими миграциями из Северо-Западной Африки. Пустыня Сахара неоднократно менялась от пышных зеленых джунглей до горячих и засушливых пустынь в последние 20 000 лет. Засушивание климата в конце ледникового периода 13000 лет назад и потепление климата привели тропические муссоны вновь с 10000 до 7000 лет до настоящего времени. Опустынивание, происходящее сегодня началось около 4200 до нашей эры. Это серьезные преобразования окружающей среды, безусловно были сильным воздействием на коренные (E1b1b) линии, в результате чего население мигрировало в эпоху неолита, и побуждение к миграциям в мягкий климат было. Было бы также неудивительно, что из Северной Африки люди пересекали Средиземное море в конце неолита. Области, наиболее пострадавшие от опустынивания были бы в районе современной Ливии. Северный Магриб находится под защитой гор, которые остановливали наступление пустыни. Египетский Нил и его дельта. Одна из гипотез заявляет, что неолитическое население Ливии мигрировало к современным Южной Италии, Греции, Македонии и Албании, и принесло с собой E-V13 линии, которая все еще находится в Ливии сегодня, а также в Иберии, Египте и Леванте , но гораздо чаще вокруг Греции. Кроме того, вместо пересечения непосредственно Средиземного моря из Туниса до Сицилии, затем в Италию и на юг Балкан, миграции могли иметь место на средиземноморском побережье, через Египет, Левант и Анатолию и, наконец, в Грецию. Некоторые мигранты могли бы принимать на запад пути к Иберии, что объясняет, почему E-V13 находится в западной Иберии, наряду с магрибскими E-M81, в то время как греки никогда не колонизировали этот регион.Распределение гаплогрупп E1b1b в странах Европы, Ближнего Востока и Северной Африки
Субклады гаплогруппы E1b1b1
E1b1b1a (или E-M78, ранее E3b1a) является наиболее распространенным среди различных субкладов гаплогруппы E среди европейцев и на Ближнем Востоке. E-M78, как полагают, мигрировали из Египта в эпоху мезолита и неолита до колонизации Ближнего Востока, где она смешивается с коренными жителями принадлежащими гаплогруппам J и G.Финикийцы, из Леванта, также способствовали распространению E1b1b1a (равно как и J2, Q и T) в таких странах, как Кипр, Мальта, Сардиния, Ибица и южная Иберия. Низкий уровень E1b1b1a в Сирии и Анатолии почти наверняка связан с конкуренцией со стороны других основных гаплогрупп неолита: G2 и J2.E-M78 делится на 4 основных ветви: E1b1b1a1 (E-V12) E1b1b1a2 (E-V13) E1b1b1a3 (E-V22) и E1b1b1a4 (E-V65), каждая в свою очередь подразделяются на "а" и "б" подсубклады .E-V13 является одним из основных маркеров неолита и сельскохозяйственных миграций из Леванта. Как и все другие субклады из E1b1b1a, E-V13 возник в Северо-Восточной Африке примерно в конце последнего ледникового периода. Его частота составляет в настоящее время гораздо выше, в Греции, Южной Италии и на Балканах, чем где-либо в связи с эффектом основателя, т.е. миграции были у небольшой группы поселенцев в основном этой линии (а также небольшое количество других Северо-Восточной африканских линий, в частности E-M123 и T). Археологические данные показывают, что область Фессалии, на севере Греции, была отправной точкой (ок. 6000 до н.э.) для распространения сельского хозяйства через Балканы и бассейн Дуная, в северной Франции на западе и Россией на востоке. Современное распределение E-V13 намекает на сильную корреляцию с неолитом и энеолитом Старой Европы, такими как Культура Винча, Боян, Марица и Караново. Тем не менее, генетические тесты трех мужчин образцы культуры LBK только показал наличие гаплогруппы F и G2a. Пример, очевидно, слишком малы, чтобы исключить, что E1b1b также попал в Европу в эпоху неолита.E-V13 также связано с древнегреческим расширением и колонизацией. За пределами Балкан и Центральной Европы, особенно часто встречается в Южной Италии, на Кипре и Южной Франции, все части классического древнегреческого мира. E-V22 является преобладающим субклад в Леванте и, следовательно, связан с финикийцами и евреями, в дополнение к распространению земледелия. Финикийцы могли бы распространить E-V22 на Сицилии, Сардинии, на юге Испании и Магриба, а евреи в Грецию и на материке Италии и Испании. Тем не менее, средиземноморский маршрут для распространения сельского хозяйства проходил через те же регионы. Поэтому невозможно узнать, в настоящее время, какой из двух периодов (неолита или античности) сыграл значительную роль в распространении V22 по всему Средиземноморью.E-V12 является самым распространенным из субкладов M78 в Египте. Его низкое присутствие в Греции и Анатолии, показывают, что он, вероятно, уже существовал, когда E переехали туда в начале неолита.E-V65 находится в Северной Африке, с максимальной частотой в Ливии и Марокко. Кроме того, вероятно, возник в Египте. В Европе она находится на низких частотах в Греции и Сицилии, но интересно, что составляет одну четвертую сардинского E. Это может быть связано с миграцией из финикийских колоний в странах Магриба и Сардинии (в Сардинии гаплогруппа I2a1 также присутствует на низких частотах, а также на побережье Алжира и Туниса, подтверждающие обмен населением между двумя регионами, может быть, когда обе территории были финикийскими колониями).E1b1b1b (E-M81, ранее E3b1b) характерно для берберов Северо-Западной Африки. В некоторых районах Марокко E1b1b1b может достичь своего пика в 80% населения. Этот субклад также находится в Иберии, Италии и южной Франции, с наибольшей концентрацией на юге Португалии (12%) и снижается по мере продвижения на север.E1b1b1c (E-M123) и ее основные ветви E1b1b1c1 (E-M34) также связаны с распространением сельского хозяйства и древними ближневосточными цивилизациями. Эта гаплогруппа достигает пика в южном Леванте (10-12% в Палестине и Ливане), откуда он расширяется во все стороны на Ближнем Востоке, Северной Африке, Южной Азии и Юго-Восточной Европы. Распределение E-M123 почти полностью совпадает с расширением сельского хозяйства (см. карту ниже), в период неолита. E-M123, кажется, идут рука об руку с гаплогруппой G2a, с той лишь разницей, что G2a достигает максимальной частоты примерно на Кавказе и в Анатолии, где крупный рогатый скот, свиньи и козы были впервые одомашнены. Внутри Европы, E-M123 более или менее распространен субклад E-V13, с высокой частотой (от 1 до 5%) наблюдается в Греции, Южной Италии, на Балканах и в бассейне Дуная, но затухание по отношению к Германии, Польши, Украине и России, где его частота составляет менее 1%.
Гаплогруппа T
Гаплогруппа T является редкой гаплогруппой в Европе. Она составляет 1% от населения на большей части континента, за исключением Греции, Македонии и Италии, где она превышает 4%, а в Иберии, где она достигает 2,5%, достигнув 10% в Кадисе и более 15% на Ибице. Максимальная частота по всему миру для гаплогруппы T наблюдается в Восточной Африке (Эритрея, Эфиопия, Сомали, Кения, Танзания) и на Ближнем Востоке (особенно на Кавказе, Южном Ираке, Юго-Западного Ирана, Омана и в Южном Египте), где на его долю приходится примерно от 5 до 15% мужчин линий. Кроме того, в этих регионах и в Европе, T находится в изолированном положении по отношению к Центральной Азии, Индии, Камеруну, Замбии и Южной Африке. Самая высокая плотность на самом деле среди фульбе народов Камеруна (18% населения).Гаплогруппа T возникла по крайней мере, 30000 лет назад, что делает её одной из старейших гаплогрупп в Евразии, чем можно объяснить огромный разрыв вокруг Африки и Южной Азии. Это также делает место её происхождения неопределенным. Современное распространение гаплогруппы T в Европе сильно коррелирует с колонизацией неолита на континенте от ближневосточных земледельцев, в которые также вошли носители гаплогрупп E1b1b, G2a, J1 и J2. Относительный максимум Эстонии весьма вероятно, связан с эффектом основателя в популяции неолита.Несмотря на то, гаплогруппа Т является более распространенной сегодня в Восточной Африке, чем где-либо еще, её связь с развитием сельского хозяйства на Ближнем Востоке является сильным аргументом в пользу Ближне-Восточного происхождения и колонизации Восточной Африки фермерами Ближнего Востока. Еще один аргумент в этом смысле указывает, что T ведет свое происхождение от гаплогруппы K, которая сама по себе отсутствует в Африке и породила большую часть евразийских гаплогрупп (L, N, O, P, Q, R и T), которые, как считается, имеют общую родину происхождения в Центральной Азии. Сильное падение Т от Кавказа, а также центральной и южной части Ирана, указывает, что первые фермеры могли бы прийти из Закавказья на юг Месопотамии и юго-запад Ирана. T поэтому может быть связана с древними шумерами и эламитами.Выше средней частота гаплогруппы T в таких местах, как Кипр, Сицилия, Тунис, Ибица, Андалусия и северная оконечность Марокко предполагают, что некоторые из гаплогруппы T можно отнести к колонизации финикийцами (1200-800 до н.э.), и что древняя Финикия возможно имела более высокую частоту T, чем Ливан сегодня.Тестирование ДНК Томаса Джефферсона показало, что президент США принадлежал к гаплогруппе Т.Продолжение
Скачать подробное филогенетическое дерево Y гаплогруппСкачать карту путей миграции Y-гаплогрупп

 Гаплогруппы чечено-ингушей: J2,J1, F, L3
Гаплогруппы чечено-ингушей: J2,J1, F, L3





























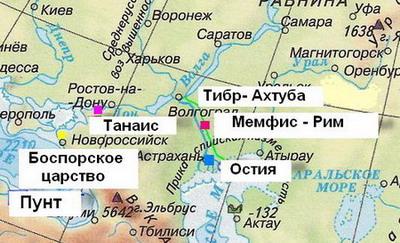
 Гаплогруппа G определяется мутацией в M201. Первый человек,с мутацией M201 , как полагают, появился около 30.000 лет назад (1200 ~ поколений), вероятно в Закавказье, но, возможно, также вдоль восточной границы на Ближнем Востоке, возможно, и на Памире. У него было относительно мало потомков.
Гаплогруппа G определяется мутацией в M201. Первый человек,с мутацией M201 , как полагают, появился около 30.000 лет назад (1200 ~ поколений), вероятно в Закавказье, но, возможно, также вдоль восточной границы на Ближнем Востоке, возможно, и на Памире. У него было относительно мало потомков.  Но разбираться особенно и не надо, потому что у всех мужчин на Земле, африканцев и неафриканцев, во всяком случае, у всех, кто делал тест на ДНК, снип-цепочки Y-хромосомы по древности одинаковы. У всех эти цепочки снипов тянутся от общих предков с шимпанзе, орангутангом, гориллой, макакой и другими древними общими с нами предками. Ниже – характерный пример, процитированный еще в 2012 году в журнале Advances in Anthropology (Klyosov, Rozhanskii, Ryabchenko):
Но разбираться особенно и не надо, потому что у всех мужчин на Земле, африканцев и неафриканцев, во всяком случае, у всех, кто делал тест на ДНК, снип-цепочки Y-хромосомы по древности одинаковы. У всех эти цепочки снипов тянутся от общих предков с шимпанзе, орангутангом, гориллой, макакой и другими древними общими с нами предками. Ниже – характерный пример, процитированный еще в 2012 году в журнале Advances in Anthropology (Klyosov, Rozhanskii, Ryabchenko):  Сравнение фрагмента нуклеотидной последовательности Y-хромосомы
человека, шимпанзе, гориллы, орангутанга и макаки Видно, что в 90% случаев нуклеотидные последовательности показанных фагментов Y-хромосомы совпадают. А у шимпанзе и человека совпадают 93 нуклеотида из 97, то есть в 96%. Так что у всех нас ДНК-корни древние, и африканцы в этом отношении никак не выделяются. Хорошо, а насколько древние? Антропологи оценивают время расхождения линии будущего человека от линии будущего шимпанзе в 4-6 миллионов лет назад, это же повторяют и генетики, хотя поиски статей с конкретными расчетами результатов не дали. А вот результаты расчета с помощью автоматического калькулятора Килина-Клёсова (ККК, 2015), иллюстрация увеличивается по клику:
Сравнение фрагмента нуклеотидной последовательности Y-хромосомы
человека, шимпанзе, гориллы, орангутанга и макаки Видно, что в 90% случаев нуклеотидные последовательности показанных фагментов Y-хромосомы совпадают. А у шимпанзе и человека совпадают 93 нуклеотида из 97, то есть в 96%. Так что у всех нас ДНК-корни древние, и африканцы в этом отношении никак не выделяются. Хорошо, а насколько древние? Антропологи оценивают время расхождения линии будущего человека от линии будущего шимпанзе в 4-6 миллионов лет назад, это же повторяют и генетики, хотя поиски статей с конкретными расчетами результатов не дали. А вот результаты расчета с помощью автоматического калькулятора Килина-Клёсова (ККК, 2015), иллюстрация увеличивается по клику:  Общий предок человека и шимпанзе жил, если округлить расчеты, 4.2±1.5 миллионов лет назад. 16-маркерный гаплотип шимпанзе был извлечен из геномной базы данных, как описано в той статье в Advances in Anthropology (2012), а в качестве обобщенного гаплотипа человека брался экстраполированный предковый гаплотип по всем неафриканским гаплогруппам, пересчитанный на времена 100 тысяч лет назад. Впрочем, можно было и не пересчитывать, вот – время до общего предка современных носителей гаплогруппы А00 и шимпанзе, иллюстрация увеличивается по клику:
Общий предок человека и шимпанзе жил, если округлить расчеты, 4.2±1.5 миллионов лет назад. 16-маркерный гаплотип шимпанзе был извлечен из геномной базы данных, как описано в той статье в Advances in Anthropology (2012), а в качестве обобщенного гаплотипа человека брался экстраполированный предковый гаплотип по всем неафриканским гаплогруппам, пересчитанный на времена 100 тысяч лет назад. Впрочем, можно было и не пересчитывать, вот – время до общего предка современных носителей гаплогруппы А00 и шимпанзе, иллюстрация увеличивается по клику:  Оно равно 4.4±1.4 миллионов лет назад, что практически одинаково в пределах погрешности расчетов. Те же времена дают все основные гаплогруппы серии А, например: • А0а-V150 имеет общего предка с шимпанзе 4.6±1.7 млн лет назад; • A1b1-M32 (Чад и Турция) 4.3±1.4 млн лет назад; • A1b1-M32, M13 (Британия) 4.2±1.4 млн лет назад, а также • гаплогруппа В, 4.4±1.4 млн лет назад. Короче, в отношении «древних ДНК-линий» африканцы ничем от неафриканцев не отличаются. И те, и другие тянут свои ДНК-линии от общего предка с шимпанзе. И у тех, и у других цепочки снипов уходят на глубины в сотни тысяч и миллионы лет. К этому мы еще вернемся. Может, под «древними африканскими ДНК-линиями» попгенетики и прочие генетики подразумевают древние времена жизни их общих предков, и полагают, что они намного более древние, чем у неафриканцев? Придется их огорчить, ситуация противоположная. Африканские предки группы А (популяций, субкладов) почти все совсем недавние. Возьмем, например, гаплогруппу А00, которая среди гаплогрупп считается самой древней. Да, для нее цепочка снипов насчитывает – по данным, приводимым YFull – 1684 снипов, что при 144 годах на снип (это число приводит YFull) дает 242500 лет от начала гаплогруппы. Действительно, YFull приводят близкий «возраст» А00, 235 тысяч лет. Но откуда эта цепочка снипов ведет – никто не знает. Может, из Африки, может, с Кавказа, может, из Сибири, может, из Европы – неизвестно. Почему-то считается, что именно из Африки. Но это опять «по понятиям», правда, откуда появившимся, непонятно, извините за игру слов. Древнейшие ДНК находили в Европе, на Русской равнине, в Сибири, а в Африке ни разу, правда, и исследований таких не было. Так откуда эти «понятия» появились? А так, стиль попгенетиков. Кстати, о возрасте гаплогруппы А00. Калькулятор КК дает для базовых гаплотипов А0 и А00 возраст общего предка (с округлением) 217000±87000 лет. Это – возраст А00 или немного старше. Так что датировка по снипам воспроизводится по гаплотипам в пределах погрешности расчетов (иллюстрация ниже увеличивается по клику).
Оно равно 4.4±1.4 миллионов лет назад, что практически одинаково в пределах погрешности расчетов. Те же времена дают все основные гаплогруппы серии А, например: • А0а-V150 имеет общего предка с шимпанзе 4.6±1.7 млн лет назад; • A1b1-M32 (Чад и Турция) 4.3±1.4 млн лет назад; • A1b1-M32, M13 (Британия) 4.2±1.4 млн лет назад, а также • гаплогруппа В, 4.4±1.4 млн лет назад. Короче, в отношении «древних ДНК-линий» африканцы ничем от неафриканцев не отличаются. И те, и другие тянут свои ДНК-линии от общего предка с шимпанзе. И у тех, и у других цепочки снипов уходят на глубины в сотни тысяч и миллионы лет. К этому мы еще вернемся. Может, под «древними африканскими ДНК-линиями» попгенетики и прочие генетики подразумевают древние времена жизни их общих предков, и полагают, что они намного более древние, чем у неафриканцев? Придется их огорчить, ситуация противоположная. Африканские предки группы А (популяций, субкладов) почти все совсем недавние. Возьмем, например, гаплогруппу А00, которая среди гаплогрупп считается самой древней. Да, для нее цепочка снипов насчитывает – по данным, приводимым YFull – 1684 снипов, что при 144 годах на снип (это число приводит YFull) дает 242500 лет от начала гаплогруппы. Действительно, YFull приводят близкий «возраст» А00, 235 тысяч лет. Но откуда эта цепочка снипов ведет – никто не знает. Может, из Африки, может, с Кавказа, может, из Сибири, может, из Европы – неизвестно. Почему-то считается, что именно из Африки. Но это опять «по понятиям», правда, откуда появившимся, непонятно, извините за игру слов. Древнейшие ДНК находили в Европе, на Русской равнине, в Сибири, а в Африке ни разу, правда, и исследований таких не было. Так откуда эти «понятия» появились? А так, стиль попгенетиков. Кстати, о возрасте гаплогруппы А00. Калькулятор КК дает для базовых гаплотипов А0 и А00 возраст общего предка (с округлением) 217000±87000 лет. Это – возраст А00 или немного старше. Так что датировка по снипам воспроизводится по гаплотипам в пределах погрешности расчетов (иллюстрация ниже увеличивается по клику).  Так вот, об общих предках африканских популяций, субкладов группы А. Они почти все по времени очень мелкие, часто всего несколько сотен лет. Иногда – между 1000 и 2000 лет. Самые древние – примерно 5000-7000 лет. И опять – где эти предки жили, никто не знает. Взглянем на гаплогруппу А00. В литературе (Mendez et al., 2013) описаны 11 гаплотипов в 95-маркерном формате (без 16 мультимаркерных гаплотипов) носителей гаплогруппы А00, все из племени Mbo из Камеруна, и еще один – афроамериканец. Расчетов в статье, конечно, нет. Все 11 гаплотипов содержат 29 мутаций, что дает 29/11/0.161 = 16 условных поколений, но с учетом того, что 45 аллелей (из 1045 аллелей) авторы определить не смогли, то до общего предка на самом деле 17 условных поколений, то есть 425±90 лет. Итак, сама гаплогруппа А00 образовалась более 200 тысяч лет назад, причем неизвестно где, а общий предок носителей А00 из Камеруна жил примерно 425 лет назад. Если добавить к серии из Камеруна еще гаплотип афроамериканца той же гаплогруппы, который отличается от предкового гаплотипа камерунцев на 11 мутаций, то есть на 11/0.161 = 68 → 73 условных поколения, то есть на 1825 лет, то их общий предок, камерунцев и афроамериканца, жил примерно (425+1825)/2 = 1125 лет назад. Примеров недавних общих предков разных мужских популяций гаплогрупп класса А, которые сейчас живут в Африке, можно привести немало. В большинстве этих популяций гаплотипы или одинаковы или почти одинаковы, даже для протяженных гаплотипов. С доступными гаплотипами гаплогруппы А00 мы уже разобрались, им четыре сотни лет. Взглянем теперь на Проект FTDNA гаплогруппы А: • В гаплогруппе/субкладе А0-V150 имеются два 25-маркерных гаплотипа, оба идентичны друг другу, то есть их общий предок жил не более 300-400 лет назад. Других гаплотипов в 25-маркерном формате или выше нет. • В гаплогруппе A0a-L981 (L994, V148, V166) между двумя доступными 67-маркерными гаплотипами есть 8 мутаций, что помещает их общего предка на 900 лет назад. • В гаплогруппе A0b-V166, L896 шесть 37-маркерных гаплотипов имеют общего предка 475±160 лет назад. • В гаплогруппе A1a-M31 («европейский кластер») пять 37-маркерных гаплотипов имеют общего предка 1175±290 лет назад. • В гаплогруппе A1a-M31 (все неизвестного происхождения) пять 37-маркерных гаплотипов имеют общего предка 1880±390 лет назад. • В гаплогруппе A1a-M31 (из 18 гаплотипов – 14 неизвестного происхождения, два из Англии, один из Швейцарии, один Кейп Верде) все 37-маркерные, все имеют общего предка 730±130 лет назад. • В гаплогруппе A1a-M31 («британский кластер») 19 гаплотипов в 67-маркерном формате имеют общего предка 740±120 лет назад. В итоге, 50 гаплотипов в 37-маркерном формате четырех предшествующих серий (А0а-М31) имеют общего предка 9270±970 лет назад. Но надо обратить внимание, что это четыре серии гаплотипов, разбросанные по разным континентам. Это – не одна «ДНК-линия», это – четыре разных ДНК-линии, с четырьмя общими предками, каждый «глубиной» от 700 до 1900 лет. Ну и где, позволительно спросить, «древнейшие ДНК-линии в Африке»? Это – миф, ничем не обоснованный. Ископаемых ДНК нет, древних линий, как мы видим, нет, к тому же, в Африке вовсе не гаплотипы группы А явяются доминирующими. Доминирует гаплогруппа E1b; в Камеруне и Чаде, и отчасти в Нигерии, доминирует гаплогруппа R1b, поэтому неудивительно, что в Африке наблюдается «высокое генетическое разнообразие», только это не от древности, а от обилия носителей разных гаплогрупп, прибывших в Африку в разные времена. Да и вообще, как можно серьезно обращать внимание на «генетическое разнобразие» в открытых системах, какой является Африка? Как я уже не раз отмечал, самым «генетически разнообразным» городом мира является, наверное, Нью-Йорк, но вряд ли кто будет серьезно утверждать, что Нью-Йорк – древнейший город мира. Кроме, наверное, популяционных генетиков, с их примитивными и бездумными критериями. Откуда же появился этот миф о «древности африканских линий»? Датировок ведь их до последнего времени не было, да и датировки бывают, как мы убедились, нескольких видов – цепочек мутаций, уходящих далеко вглубь, до приматов; цепочек мутаций, ведущих ко временам образования соответствующих гаплогрупп; и датировок общих предков современных носителей соответствующих гаплогрупп и субкладов. Ни одна из этих датировок не «укоренена» территориально, то есть мы не знаем, на какой территории, на каком континенте образовалась та или иная гаплогруппа. Мы можем только набирать сведения и складывать фрагменты общей картины, но Африка в этом отношении находится на одном из последних мест по накопленным знаниям. Но фантазий, натяжек и откровенных фальсификаций про них со стороны попгенетиков – хоть отбавляй. Собственно, это характерно для популяционных генетиков, но они уроки из принципиальных ошибок не извлекают. Еще не так давно они утверждали, что гаплогруппа R1b образовалась в Европе 30 тысяч лет назад, и сначала приняли в штыки, а потом напрочь игнорировали сведения и датировки ДНК-генеалогии, что R1b прибыла из Южной Сибири длинным миграционным путем в Европу примерно 4800 лет назад (Klyosov, 2009; Klyosov, 2012). Называли это «псевдонаукой», особенно в этом усердствовали попгенетики Балановские. Разворот произошел в 2013 году, когда неподалеку от Байкала нашли ископаемую гаплогруппу R с археологической датировкой 24 тысячи лет назад. Лед тронулся, и попгенетики, не моргнув глазом, не повинившись и не цитируя статьи с датировками ДНК-генеалогии, стали повторять то, что уже было опубликовано нами несколько лет назад. Например, в недавней статье Batini, Jobling и других (2015) датировки гаплогруппы R1b в Европе уже даются как R1b-L11 4510 лет (доверительный интервал 3920-5160 лет назад) и R1b-P312 4210 лет (доверительный интервал 3700-4790 лет назад). YFull дают датировки R1b-U106 4900 лет (доверительный интервал 4400-5400 лет назад), R1b-P312 4600 лет (для TMRCA, доверительный интервал 4300-5000 лет назад). К сожалению, приходится повторять, что попгенетики уроков из своих ошибок не извлекают. Ситуация повторяется с Африкой. Данных нет, или данные говорят о другом, но их придумывают и искажают. Зачем это им нужно? Для нормального человека звучит невероятно, но это ментальность вассалов, которые лебезят перед сюзереном. Сюзерен решает, кого пригреть, кого пригласить на конференцию, кому дать грант, кого взять в коллективную статью, ввести в комиссию или комитет, в общем, в инфраструктуру. Без сюзерена вассал – никто, и вассал это прекрасно понимает. А зачем это нужно сюзерену? О, там большая политика. Вассала к этим вопросам не допускают. Вассал знает, что если он публично усомнится в африканском происхождении человечества, то его дни сочтены. Поэтому он, вассал, готов разорвать на части любого, кто сомневается или выставляет данные против. А поскольку он разорвать на части не может, то цель – найти компромат, состряпать ложь, передернуть, исказить – других способов или аргументов у вассала нет, во всяком случае, научных. Посмотрим на схему гаплогрупп и субкладов группы А, приведенную согласно классификации ISOGG, в сокращенном виде:
Так вот, об общих предках африканских популяций, субкладов группы А. Они почти все по времени очень мелкие, часто всего несколько сотен лет. Иногда – между 1000 и 2000 лет. Самые древние – примерно 5000-7000 лет. И опять – где эти предки жили, никто не знает. Взглянем на гаплогруппу А00. В литературе (Mendez et al., 2013) описаны 11 гаплотипов в 95-маркерном формате (без 16 мультимаркерных гаплотипов) носителей гаплогруппы А00, все из племени Mbo из Камеруна, и еще один – афроамериканец. Расчетов в статье, конечно, нет. Все 11 гаплотипов содержат 29 мутаций, что дает 29/11/0.161 = 16 условных поколений, но с учетом того, что 45 аллелей (из 1045 аллелей) авторы определить не смогли, то до общего предка на самом деле 17 условных поколений, то есть 425±90 лет. Итак, сама гаплогруппа А00 образовалась более 200 тысяч лет назад, причем неизвестно где, а общий предок носителей А00 из Камеруна жил примерно 425 лет назад. Если добавить к серии из Камеруна еще гаплотип афроамериканца той же гаплогруппы, который отличается от предкового гаплотипа камерунцев на 11 мутаций, то есть на 11/0.161 = 68 → 73 условных поколения, то есть на 1825 лет, то их общий предок, камерунцев и афроамериканца, жил примерно (425+1825)/2 = 1125 лет назад. Примеров недавних общих предков разных мужских популяций гаплогрупп класса А, которые сейчас живут в Африке, можно привести немало. В большинстве этих популяций гаплотипы или одинаковы или почти одинаковы, даже для протяженных гаплотипов. С доступными гаплотипами гаплогруппы А00 мы уже разобрались, им четыре сотни лет. Взглянем теперь на Проект FTDNA гаплогруппы А: • В гаплогруппе/субкладе А0-V150 имеются два 25-маркерных гаплотипа, оба идентичны друг другу, то есть их общий предок жил не более 300-400 лет назад. Других гаплотипов в 25-маркерном формате или выше нет. • В гаплогруппе A0a-L981 (L994, V148, V166) между двумя доступными 67-маркерными гаплотипами есть 8 мутаций, что помещает их общего предка на 900 лет назад. • В гаплогруппе A0b-V166, L896 шесть 37-маркерных гаплотипов имеют общего предка 475±160 лет назад. • В гаплогруппе A1a-M31 («европейский кластер») пять 37-маркерных гаплотипов имеют общего предка 1175±290 лет назад. • В гаплогруппе A1a-M31 (все неизвестного происхождения) пять 37-маркерных гаплотипов имеют общего предка 1880±390 лет назад. • В гаплогруппе A1a-M31 (из 18 гаплотипов – 14 неизвестного происхождения, два из Англии, один из Швейцарии, один Кейп Верде) все 37-маркерные, все имеют общего предка 730±130 лет назад. • В гаплогруппе A1a-M31 («британский кластер») 19 гаплотипов в 67-маркерном формате имеют общего предка 740±120 лет назад. В итоге, 50 гаплотипов в 37-маркерном формате четырех предшествующих серий (А0а-М31) имеют общего предка 9270±970 лет назад. Но надо обратить внимание, что это четыре серии гаплотипов, разбросанные по разным континентам. Это – не одна «ДНК-линия», это – четыре разных ДНК-линии, с четырьмя общими предками, каждый «глубиной» от 700 до 1900 лет. Ну и где, позволительно спросить, «древнейшие ДНК-линии в Африке»? Это – миф, ничем не обоснованный. Ископаемых ДНК нет, древних линий, как мы видим, нет, к тому же, в Африке вовсе не гаплотипы группы А явяются доминирующими. Доминирует гаплогруппа E1b; в Камеруне и Чаде, и отчасти в Нигерии, доминирует гаплогруппа R1b, поэтому неудивительно, что в Африке наблюдается «высокое генетическое разнообразие», только это не от древности, а от обилия носителей разных гаплогрупп, прибывших в Африку в разные времена. Да и вообще, как можно серьезно обращать внимание на «генетическое разнобразие» в открытых системах, какой является Африка? Как я уже не раз отмечал, самым «генетически разнообразным» городом мира является, наверное, Нью-Йорк, но вряд ли кто будет серьезно утверждать, что Нью-Йорк – древнейший город мира. Кроме, наверное, популяционных генетиков, с их примитивными и бездумными критериями. Откуда же появился этот миф о «древности африканских линий»? Датировок ведь их до последнего времени не было, да и датировки бывают, как мы убедились, нескольких видов – цепочек мутаций, уходящих далеко вглубь, до приматов; цепочек мутаций, ведущих ко временам образования соответствующих гаплогрупп; и датировок общих предков современных носителей соответствующих гаплогрупп и субкладов. Ни одна из этих датировок не «укоренена» территориально, то есть мы не знаем, на какой территории, на каком континенте образовалась та или иная гаплогруппа. Мы можем только набирать сведения и складывать фрагменты общей картины, но Африка в этом отношении находится на одном из последних мест по накопленным знаниям. Но фантазий, натяжек и откровенных фальсификаций про них со стороны попгенетиков – хоть отбавляй. Собственно, это характерно для популяционных генетиков, но они уроки из принципиальных ошибок не извлекают. Еще не так давно они утверждали, что гаплогруппа R1b образовалась в Европе 30 тысяч лет назад, и сначала приняли в штыки, а потом напрочь игнорировали сведения и датировки ДНК-генеалогии, что R1b прибыла из Южной Сибири длинным миграционным путем в Европу примерно 4800 лет назад (Klyosov, 2009; Klyosov, 2012). Называли это «псевдонаукой», особенно в этом усердствовали попгенетики Балановские. Разворот произошел в 2013 году, когда неподалеку от Байкала нашли ископаемую гаплогруппу R с археологической датировкой 24 тысячи лет назад. Лед тронулся, и попгенетики, не моргнув глазом, не повинившись и не цитируя статьи с датировками ДНК-генеалогии, стали повторять то, что уже было опубликовано нами несколько лет назад. Например, в недавней статье Batini, Jobling и других (2015) датировки гаплогруппы R1b в Европе уже даются как R1b-L11 4510 лет (доверительный интервал 3920-5160 лет назад) и R1b-P312 4210 лет (доверительный интервал 3700-4790 лет назад). YFull дают датировки R1b-U106 4900 лет (доверительный интервал 4400-5400 лет назад), R1b-P312 4600 лет (для TMRCA, доверительный интервал 4300-5000 лет назад). К сожалению, приходится повторять, что попгенетики уроков из своих ошибок не извлекают. Ситуация повторяется с Африкой. Данных нет, или данные говорят о другом, но их придумывают и искажают. Зачем это им нужно? Для нормального человека звучит невероятно, но это ментальность вассалов, которые лебезят перед сюзереном. Сюзерен решает, кого пригреть, кого пригласить на конференцию, кому дать грант, кого взять в коллективную статью, ввести в комиссию или комитет, в общем, в инфраструктуру. Без сюзерена вассал – никто, и вассал это прекрасно понимает. А зачем это нужно сюзерену? О, там большая политика. Вассала к этим вопросам не допускают. Вассал знает, что если он публично усомнится в африканском происхождении человечества, то его дни сочтены. Поэтому он, вассал, готов разорвать на части любого, кто сомневается или выставляет данные против. А поскольку он разорвать на части не может, то цель – найти компромат, состряпать ложь, передернуть, исказить – других способов или аргументов у вассала нет, во всяком случае, научных. Посмотрим на схему гаплогрупп и субкладов группы А, приведенную согласно классификации ISOGG, в сокращенном виде:  В этой схеме – источник принципиальной путаницы попенетиков. Они не смогли увидеть и осознать, что эта лесенка субкладов представляет собой серию вилок, которые периодически расходятся на «африканские» (по сегодняшнему месту жительства) и неафриканские ветви. Опишем кратко эти вилки, а потом рассмотрим их подробнее – когда они образовались, и кто являются потомками субкладов по обе стороны тех вилок. Итак, вилок всего четыре. Первой от ствола, уходящего вглубь эволюционного дерева человечества (точнее, его Y-хромосомы), отходит вилка, знаменующая, видимо, начало Homo sapiens – расхождение на А00 и А0-Т. Вторая – расхождение А0-Т на А0 и А1. Третья – расхождение А1 на А1a и A1b. Четвертая – расхождение A1b на A1b1 и ВТ. Если упрощенно – то каждая вилка расходится на африканскую (по нынешнему месту жительства) и неафриканскую стороны. А00, А0, A1a и A1b1 уходят в Африку, противоположные стороны вилки A0-T, A1, A1b и BT остаются вне Африки. «Упрощенно» – потому что вилка на А00 и A0-T образовалась еще до расхождения на африканцев и неафриканцев (это расхождение произошло примерно 160 тысяч лет назад), а впоследствии многие неафриканцы перебирались в Африку – это и многочисленные носители гаплогрупп E1b, J2, R1b, и так далее. Среди них – миллионы современных жителей Западной Африки (берберы, жители Камеруна и Чада), египтяне, потомки буров в Южной Африке, и так далее. Немало жителей Европы и Ближнего Востока имеют гаплогруппу A1a, как недавно показывал И.Л. Рожанский в статье на Переформате. Да и вообще носителей группы А в Африке относительно мало – часто на уровне единиц процента или того меньше. Рассмотрим вилки подробнее. Первая вилка – расхождение предшественника Homo sapiens (или Homo sapiens в начальной стадии, примерно 230 тысяч лет назад) на гаплогруппы А00 и А0-Т. Нисходящих субкладов от А00 пока не обнаружено, и ее нынешние носители обнаружены только в Африке в количестве 11 человек (см. выше), с общим предком несколько сотен лет назад, а также несколько афроамериканцев с одинаковыми гаплотипами. Снипы гаплогруппы А0-Т, напротив, имеются у всех неафриканцев, в том числе у всех нас, читателей Переформата, хотя только у мужчин. И, разумеется, у всех африканцев, носителей гаплогрупп E1b, J2, R1b, В и других, которые прибыли в Африку относительно недавно. Но помимо них снипы группы А0-Т должны быть и у всех африканцев, носителей групп А, потому эти снипы предшествуют разделению африканских и неафриканских линий (примерно 160 тысяч лет назад). Впрочем, речь идет о промежуточных снипах в цепочках наших и африканских снипов, носителей А0-Т как терминальных снипов пока ни у кого не выявлено, ни в Африке, ни вне Африки. Поэтому называть его «африканским» никакого смысла не имеет. К тому, что у всех нас, мужчин, есть множество снипов гаплогруппы А0-Т – ниже приведен список части из 587 снипов этой гаплогруппы, из которых у меня есть L1090, L1121, L1124, L1125, L1127, L1235 и многие другие: Мой список снипов BigY CTS10042, CTS10627, CTS10834, CTS10847, CTS10976, CTS10993, CTS11148, CTS11184, CTS11411, CTS11468, CTS11530, CTS1169, CTS11720, CTS11734, CTS11853, CTS11913, CTS11948, CTS12010, CTS1211, CTS1277, CTS1278, CTS12947, CTS12972, CTS1340, CTS1367, CTS1415, CTS1416, CTS1417, CTS1419, CTS1422, CTS1619, CTS1738, CTS1795, CTS1907, CTS2134, CTS262, CTS2626, CTS2636, CTS2637, CTS2638, CTS2639, CTS2663, CTS 2680, CTS2891, CTS2988, CTS2992, CTS3067, CTS3075, CTS3229, CTS3230, CTS3231, CTS3316, CTS3321, CTS3358, CTS3402, CTS3441, CTS3527, CTS3534, CTS3548, CTS3551, CTS358, CTS3607, CTS3622, CTS3649, CTS3650, CTS3654, CTS3697, CTS3775, CTS3848, CTS3943, CTS3984, CTS4259, CTS4465, CTS4862, CTS501, CTS5287, CTS5423, CTS5454, CTS5508, CTS5577, CTS5648, CTS570, CTS5936, CTS5979, CTS623, CTS6918, CTS6985, CTS7191, CTS7275, CTS7278, CTS7400, CTS7500, CTS7650, CTS7659, CTS7941, CTS8008, CTS8127, CTS8350, CTS8507, CTS8508, CTS8591, CTS8626, CTS8665, CTS8797, CTS8816, CTS8896, CTS9018, CTS903, CTS9108, CTS9510, CTS9515, CTS9539, CTS9596, CTS9733, CTS9739, CTS9754, CTS9779, F1050, F1209, F1224, F132, F1586, F184, F185, F2957, F3032, F313, F3159, F3166, F3185, F3194, F3197, F3337, F3561, F3564, F3644, F3650, F3692, F4099, F686, F719, F773, F82, F937, F989, IMS-JST002612, IMS-ST022448_3, IMS-ST022448_6, IMS-ST022449_3, IMS-ST022449_6, IMS-ST022451_2, K511, L1002, L1004, L1005, L1009, L1013, L1028, L104, L105, L1053, L1060, L1061, L1062, L1067, L1071, L108, L1089, L1090, L1095, L1098, L1099, L110, L1101, L1105, L1112, L1116, L1118, L112, L1120, L1121, L1123, L1124, L1125, L1127, L1129, L113, L1132, L1135, L1136, L1137, L114, L1142, L1143, L1145, L1150, L1155, L1179, L1185, L1120, L1225, L1235, L132, L1346, L1347, L1352, L1480, L1492, L15, L16, L168, L248, L268, L313, L350, L352, L413, L418, L438, L440, L449, L457, L468, L469, L498, L543, L58, L604, L721, L74, L741, L747, L76, L760, L768, L777, L779, L781, L819, L82, L821, L851, L875, L882, L962, L969, L970, L977, L986, L989, M168, M173, M198, M207, M213, M235, M251, M294, M299, M306, M417, M42, M420, M448, M449, M45, M459, M511, M512, M513, M514, M515, M516, M526, M74, M89, M9, M94, P108, P128, P131, P132, P133, P134, P135, P136, P138, P139, P14, P140, P141, P143, P145, P146, P148, P149, P151, P157, P158, P159, P160, P161, P163, P166, P187, P19_1, P207, P224, P225, P226, P228, P229, P230, P231, P232, P233, P234, P235, P236, P237, P238, P239, P242, P243, P244, P245, P27.1_2, P280, P281, P282, P283, P284, P285, P286, P294, P305, P316, PAGES00026, PAGES00081, PAGES00083, PF1345, PF1378, PF1389, PF1396, PF1402, PF1403, PF1405, PF1407, PF1408, PF1411, PF1412, PF1414, PF1415, PF1418, PF1419, PF2589, PF2590, PF2591, PF2608, PF2610, PF2611, PF2614, PF2615, PF2616, PF2619, PF2620, PF2621, PF2622, PF2626, PF2629, PF2631, PF2634, PF2635, PF2637, PF2640, PF2642, PF2647, PF2651, PF2653, PF2658, PF2660, PF2683, PF2685, PF288, PF2690, PF2694, PF2695, PF2700, PF2709, PF2716, PF2718, PF2722, PF2723, PF2729, PF2731, PF2732, PF2736, PF2737, PF2739, PF2742, PF2747, PF2748, PF2749, PF2752, PF2753, PF2754, PF2755, PF2756, PF2757, PF2758, PF2770, PF2772, PF2988, PF349, PF3494, PF3495, PF3496, PF3497, PF3500, PF4522, PF5459, PF5469, PF5477, PF5483, PF5488, PF5490, PF5498, PF5500, PF5503, PF5513, PF5861, PF5862, PF5863, PF5864, PF5869, PF5870, PF5871, PF5872, PF5876, PF5881, PF5882, PF5885, PF5886, PF5888, PF5889, PF5895, PF5908, PF5911, PF5914, PF5916, PF5919, PF5920, PF5936, PF5940, PF5943, PF5944, PF5945, PF5949, PF5951, PF5953, PF5954, PF5955, PF5956, PF5957, PF5958, PF5964, PF5965, PF5967, PF5970, PF5971, PF5975, PF5978, PF5980, PF5981, PF5982, PF5984, PF5985, PF5991, PF6013, PF6016, PF6047, PF6054, PF6056, PF6057, PF6066, PF6069, PF6071, PF6072, PF6073, PF6114, PF6116, PF6118, PF6121, PF6132, PF6133, PF6143, PF6153, PF6156, PF6158, PF6159, PF6160, PF6162, PF6163, PF6165, PF6168, PF6169, PF6170, PF6172, PF6179, PF6181, PF6194, PF6195, PF6201, PF6204, PF6211, PF6212, PF6213, PF6214, PF6215, PF6216, PF6217, PF6218, PF6220, PF6222, PF6230, PF6231, PF6233, PF6512, PF682, PF7514, PF7522, PF7527, PF7530, PF7531, PF7532, PF7534, PF7540, PF7542, PK1, PR1220, PR202, PR2119, PR5761, PR855, rs161492_1, V221, V238, V241, V250, Y2902, Y32, Y33, Y34, Y36, Y783.2, YSC0000057, YSC0000077, YSC0000081, YSC0000107, YSC0000108, YSC0000109, YSC0000110, YSC0000193, YSC0000350, YSC0000913, Z1331, Z280, Z282, Z5997, Z6030, Z6032. Фрагмент из списка снипов гаплогруппы A0-T (приведены 182 снипа из 587)
В этой схеме – источник принципиальной путаницы попенетиков. Они не смогли увидеть и осознать, что эта лесенка субкладов представляет собой серию вилок, которые периодически расходятся на «африканские» (по сегодняшнему месту жительства) и неафриканские ветви. Опишем кратко эти вилки, а потом рассмотрим их подробнее – когда они образовались, и кто являются потомками субкладов по обе стороны тех вилок. Итак, вилок всего четыре. Первой от ствола, уходящего вглубь эволюционного дерева человечества (точнее, его Y-хромосомы), отходит вилка, знаменующая, видимо, начало Homo sapiens – расхождение на А00 и А0-Т. Вторая – расхождение А0-Т на А0 и А1. Третья – расхождение А1 на А1a и A1b. Четвертая – расхождение A1b на A1b1 и ВТ. Если упрощенно – то каждая вилка расходится на африканскую (по нынешнему месту жительства) и неафриканскую стороны. А00, А0, A1a и A1b1 уходят в Африку, противоположные стороны вилки A0-T, A1, A1b и BT остаются вне Африки. «Упрощенно» – потому что вилка на А00 и A0-T образовалась еще до расхождения на африканцев и неафриканцев (это расхождение произошло примерно 160 тысяч лет назад), а впоследствии многие неафриканцы перебирались в Африку – это и многочисленные носители гаплогрупп E1b, J2, R1b, и так далее. Среди них – миллионы современных жителей Западной Африки (берберы, жители Камеруна и Чада), египтяне, потомки буров в Южной Африке, и так далее. Немало жителей Европы и Ближнего Востока имеют гаплогруппу A1a, как недавно показывал И.Л. Рожанский в статье на Переформате. Да и вообще носителей группы А в Африке относительно мало – часто на уровне единиц процента или того меньше. Рассмотрим вилки подробнее. Первая вилка – расхождение предшественника Homo sapiens (или Homo sapiens в начальной стадии, примерно 230 тысяч лет назад) на гаплогруппы А00 и А0-Т. Нисходящих субкладов от А00 пока не обнаружено, и ее нынешние носители обнаружены только в Африке в количестве 11 человек (см. выше), с общим предком несколько сотен лет назад, а также несколько афроамериканцев с одинаковыми гаплотипами. Снипы гаплогруппы А0-Т, напротив, имеются у всех неафриканцев, в том числе у всех нас, читателей Переформата, хотя только у мужчин. И, разумеется, у всех африканцев, носителей гаплогрупп E1b, J2, R1b, В и других, которые прибыли в Африку относительно недавно. Но помимо них снипы группы А0-Т должны быть и у всех африканцев, носителей групп А, потому эти снипы предшествуют разделению африканских и неафриканских линий (примерно 160 тысяч лет назад). Впрочем, речь идет о промежуточных снипах в цепочках наших и африканских снипов, носителей А0-Т как терминальных снипов пока ни у кого не выявлено, ни в Африке, ни вне Африки. Поэтому называть его «африканским» никакого смысла не имеет. К тому, что у всех нас, мужчин, есть множество снипов гаплогруппы А0-Т – ниже приведен список части из 587 снипов этой гаплогруппы, из которых у меня есть L1090, L1121, L1124, L1125, L1127, L1235 и многие другие: Мой список снипов BigY CTS10042, CTS10627, CTS10834, CTS10847, CTS10976, CTS10993, CTS11148, CTS11184, CTS11411, CTS11468, CTS11530, CTS1169, CTS11720, CTS11734, CTS11853, CTS11913, CTS11948, CTS12010, CTS1211, CTS1277, CTS1278, CTS12947, CTS12972, CTS1340, CTS1367, CTS1415, CTS1416, CTS1417, CTS1419, CTS1422, CTS1619, CTS1738, CTS1795, CTS1907, CTS2134, CTS262, CTS2626, CTS2636, CTS2637, CTS2638, CTS2639, CTS2663, CTS 2680, CTS2891, CTS2988, CTS2992, CTS3067, CTS3075, CTS3229, CTS3230, CTS3231, CTS3316, CTS3321, CTS3358, CTS3402, CTS3441, CTS3527, CTS3534, CTS3548, CTS3551, CTS358, CTS3607, CTS3622, CTS3649, CTS3650, CTS3654, CTS3697, CTS3775, CTS3848, CTS3943, CTS3984, CTS4259, CTS4465, CTS4862, CTS501, CTS5287, CTS5423, CTS5454, CTS5508, CTS5577, CTS5648, CTS570, CTS5936, CTS5979, CTS623, CTS6918, CTS6985, CTS7191, CTS7275, CTS7278, CTS7400, CTS7500, CTS7650, CTS7659, CTS7941, CTS8008, CTS8127, CTS8350, CTS8507, CTS8508, CTS8591, CTS8626, CTS8665, CTS8797, CTS8816, CTS8896, CTS9018, CTS903, CTS9108, CTS9510, CTS9515, CTS9539, CTS9596, CTS9733, CTS9739, CTS9754, CTS9779, F1050, F1209, F1224, F132, F1586, F184, F185, F2957, F3032, F313, F3159, F3166, F3185, F3194, F3197, F3337, F3561, F3564, F3644, F3650, F3692, F4099, F686, F719, F773, F82, F937, F989, IMS-JST002612, IMS-ST022448_3, IMS-ST022448_6, IMS-ST022449_3, IMS-ST022449_6, IMS-ST022451_2, K511, L1002, L1004, L1005, L1009, L1013, L1028, L104, L105, L1053, L1060, L1061, L1062, L1067, L1071, L108, L1089, L1090, L1095, L1098, L1099, L110, L1101, L1105, L1112, L1116, L1118, L112, L1120, L1121, L1123, L1124, L1125, L1127, L1129, L113, L1132, L1135, L1136, L1137, L114, L1142, L1143, L1145, L1150, L1155, L1179, L1185, L1120, L1225, L1235, L132, L1346, L1347, L1352, L1480, L1492, L15, L16, L168, L248, L268, L313, L350, L352, L413, L418, L438, L440, L449, L457, L468, L469, L498, L543, L58, L604, L721, L74, L741, L747, L76, L760, L768, L777, L779, L781, L819, L82, L821, L851, L875, L882, L962, L969, L970, L977, L986, L989, M168, M173, M198, M207, M213, M235, M251, M294, M299, M306, M417, M42, M420, M448, M449, M45, M459, M511, M512, M513, M514, M515, M516, M526, M74, M89, M9, M94, P108, P128, P131, P132, P133, P134, P135, P136, P138, P139, P14, P140, P141, P143, P145, P146, P148, P149, P151, P157, P158, P159, P160, P161, P163, P166, P187, P19_1, P207, P224, P225, P226, P228, P229, P230, P231, P232, P233, P234, P235, P236, P237, P238, P239, P242, P243, P244, P245, P27.1_2, P280, P281, P282, P283, P284, P285, P286, P294, P305, P316, PAGES00026, PAGES00081, PAGES00083, PF1345, PF1378, PF1389, PF1396, PF1402, PF1403, PF1405, PF1407, PF1408, PF1411, PF1412, PF1414, PF1415, PF1418, PF1419, PF2589, PF2590, PF2591, PF2608, PF2610, PF2611, PF2614, PF2615, PF2616, PF2619, PF2620, PF2621, PF2622, PF2626, PF2629, PF2631, PF2634, PF2635, PF2637, PF2640, PF2642, PF2647, PF2651, PF2653, PF2658, PF2660, PF2683, PF2685, PF288, PF2690, PF2694, PF2695, PF2700, PF2709, PF2716, PF2718, PF2722, PF2723, PF2729, PF2731, PF2732, PF2736, PF2737, PF2739, PF2742, PF2747, PF2748, PF2749, PF2752, PF2753, PF2754, PF2755, PF2756, PF2757, PF2758, PF2770, PF2772, PF2988, PF349, PF3494, PF3495, PF3496, PF3497, PF3500, PF4522, PF5459, PF5469, PF5477, PF5483, PF5488, PF5490, PF5498, PF5500, PF5503, PF5513, PF5861, PF5862, PF5863, PF5864, PF5869, PF5870, PF5871, PF5872, PF5876, PF5881, PF5882, PF5885, PF5886, PF5888, PF5889, PF5895, PF5908, PF5911, PF5914, PF5916, PF5919, PF5920, PF5936, PF5940, PF5943, PF5944, PF5945, PF5949, PF5951, PF5953, PF5954, PF5955, PF5956, PF5957, PF5958, PF5964, PF5965, PF5967, PF5970, PF5971, PF5975, PF5978, PF5980, PF5981, PF5982, PF5984, PF5985, PF5991, PF6013, PF6016, PF6047, PF6054, PF6056, PF6057, PF6066, PF6069, PF6071, PF6072, PF6073, PF6114, PF6116, PF6118, PF6121, PF6132, PF6133, PF6143, PF6153, PF6156, PF6158, PF6159, PF6160, PF6162, PF6163, PF6165, PF6168, PF6169, PF6170, PF6172, PF6179, PF6181, PF6194, PF6195, PF6201, PF6204, PF6211, PF6212, PF6213, PF6214, PF6215, PF6216, PF6217, PF6218, PF6220, PF6222, PF6230, PF6231, PF6233, PF6512, PF682, PF7514, PF7522, PF7527, PF7530, PF7531, PF7532, PF7534, PF7540, PF7542, PK1, PR1220, PR202, PR2119, PR5761, PR855, rs161492_1, V221, V238, V241, V250, Y2902, Y32, Y33, Y34, Y36, Y783.2, YSC0000057, YSC0000077, YSC0000081, YSC0000107, YSC0000108, YSC0000109, YSC0000110, YSC0000193, YSC0000350, YSC0000913, Z1331, Z280, Z282, Z5997, Z6030, Z6032. Фрагмент из списка снипов гаплогруппы A0-T (приведены 182 снипа из 587)  Понятие «вилка» не стоит понимать буквально, А00 и А0-Т могли образоваться с разницей в десятки тысяч лет. По данным YFull, до начала А00 насчитывается 1684 снипа, до начала А0-Т – 587 снипов, но возраст обеих гаплогрупп почему-то выставлен как одинаковый, 235 тысяч лет назад. На такую повторяющуюся «дихотомию» в презентации данных YFull мы уже не раз обращали внимание. В любом случае, искать у нас, читателей Переформата, снипы гаплогруппы А00 бесполезно, нет их у нас, ни одного из тех 1684 снипов. Но важно то, что А0-Т образует очередную вилку, на гаплогруппы А0 и А1. Вторая вилка – расхождение А0-Т на А0 и А1. Потомки А0 сейчас живут в Африке, потомки А1 – мы с вами, читатели Переформата. У всех нас в цепочках снипов есть снипы субклада А1 (см. ниже), но это опять не значит, что наши предки вышли из Африки. Это опять промежуточные снипы, и никого с терминальными снипами А1 пока не выявлено, ни в Африке, ни вне Африки. Поэтому называть его «африканским» никакого смысла тоже не имеет. По данным, приведенным YFull, к A0 ведет 899 снипов (это 129460 лет, если снип образуется в среднем раз в 144 года, как считали те же YFull), к А1 – 62 снипа (8900 лет), возраст А0 не выставлен, но для А1 выставлено 146300 лет «возраста». Опять загадка, как это может быть. Судя по этим данным, на протяжении 100 тысяч лет между А00 и А0-Т, гаплогрупп между ними вовсе не образовывалось, и к чему те 62 снипа относятся – непонятно. Но здесь важнее то, что А0 – это «африканская» гаплогруппа (опять по нынешнему месту жительства), а А1 – неафриканская. На схеме выше показано семь субкладов гаплогрупы А0, и снипов никаких из них у нас, читателей Переформата, нет, а снипы гаплогруппы А1 есть у нас всех (я не буду больше повторять, что это относится к обладателям Y-хромосомы, то есть к мужчинам, и что «читатель Переформата» относится к неафриканцам, потомкам гаплогруппы ВТ, которые не перебрались на жительство в Африку). Повторю, что «африканских» снипов А0 у нас нет, можно не искать, нет ни одного из 899 тех снипов. Не выходили наши предки из Африки, или, во всяком случае, не являются потомками ни А00, ни А0. Надо сказать, что в Африке носителей гаплогруппы А0 обнаружено немного. Scozzari et al (2012) указывали, что только после долгих поисков нашли трех (!) носителей гаплогруппы А0, все в Камеруне, и отмечали, что эта гаплогруппа имеет в Африке очень низкую частоту встречаемости. Ниже приведен список из 62 снипов гаплогруппы А1, из которых у меня есть L986, L989, L1002, L1009, L1112, V238, V241, V250. Остальных нет потому, что не все снипы у каждого определяются. Список снипов гаплогруппы A1
Понятие «вилка» не стоит понимать буквально, А00 и А0-Т могли образоваться с разницей в десятки тысяч лет. По данным YFull, до начала А00 насчитывается 1684 снипа, до начала А0-Т – 587 снипов, но возраст обеих гаплогрупп почему-то выставлен как одинаковый, 235 тысяч лет назад. На такую повторяющуюся «дихотомию» в презентации данных YFull мы уже не раз обращали внимание. В любом случае, искать у нас, читателей Переформата, снипы гаплогруппы А00 бесполезно, нет их у нас, ни одного из тех 1684 снипов. Но важно то, что А0-Т образует очередную вилку, на гаплогруппы А0 и А1. Вторая вилка – расхождение А0-Т на А0 и А1. Потомки А0 сейчас живут в Африке, потомки А1 – мы с вами, читатели Переформата. У всех нас в цепочках снипов есть снипы субклада А1 (см. ниже), но это опять не значит, что наши предки вышли из Африки. Это опять промежуточные снипы, и никого с терминальными снипами А1 пока не выявлено, ни в Африке, ни вне Африки. Поэтому называть его «африканским» никакого смысла тоже не имеет. По данным, приведенным YFull, к A0 ведет 899 снипов (это 129460 лет, если снип образуется в среднем раз в 144 года, как считали те же YFull), к А1 – 62 снипа (8900 лет), возраст А0 не выставлен, но для А1 выставлено 146300 лет «возраста». Опять загадка, как это может быть. Судя по этим данным, на протяжении 100 тысяч лет между А00 и А0-Т, гаплогрупп между ними вовсе не образовывалось, и к чему те 62 снипа относятся – непонятно. Но здесь важнее то, что А0 – это «африканская» гаплогруппа (опять по нынешнему месту жительства), а А1 – неафриканская. На схеме выше показано семь субкладов гаплогрупы А0, и снипов никаких из них у нас, читателей Переформата, нет, а снипы гаплогруппы А1 есть у нас всех (я не буду больше повторять, что это относится к обладателям Y-хромосомы, то есть к мужчинам, и что «читатель Переформата» относится к неафриканцам, потомкам гаплогруппы ВТ, которые не перебрались на жительство в Африку). Повторю, что «африканских» снипов А0 у нас нет, можно не искать, нет ни одного из 899 тех снипов. Не выходили наши предки из Африки, или, во всяком случае, не являются потомками ни А00, ни А0. Надо сказать, что в Африке носителей гаплогруппы А0 обнаружено немного. Scozzari et al (2012) указывали, что только после долгих поисков нашли трех (!) носителей гаплогруппы А0, все в Камеруне, и отмечали, что эта гаплогруппа имеет в Африке очень низкую частоту встречаемости. Ниже приведен список из 62 снипов гаплогруппы А1, из которых у меня есть L986, L989, L1002, L1009, L1112, V238, V241, V250. Остальных нет потому, что не все снипы у каждого определяются. Список снипов гаплогруппы A1  Третья вилка – расхождение А1 на A1a и A1b. Потомки A1a сейчас живут в Африке и по всему миру (в частности, в ряде европейских стран и на Ближнем Востоке). Снипы А1b есть у всех нас, читатели Переформата. Но это опять не значит, что наши предки вышли из Африки. Это опять промежуточные снипы, и никого с терминальными снипами А1b пока не выявлено, ни в Африке, ни вне Африки. Поэтому называть его «африканским» опять никакого смысла не имеет. По данным, приведенным YFull, к A1a ведет 495 снипов (это 71 тысяча лет, если снип образуется в среднем раз в 144 года), к А1b – 44 снипа (6300 лет), но почему-то им опять было выставлено по 132600 лет «возраста». Опять загадка, на каком основании так делается. Хотя носители гаплогруппы A1a разбросаны по всему миру (поэтому в любом случае это не «африканская» гаплогруппа, даже по такому условному критерию, как современное место жительства), их мало. И.Р. Рожанский уже приводил в своей статье «Африканцы на севере Европы. Африканцы ли?», что гаплогруппа A1a-M31 чрезвычайно редкая, и даже в Африке ее наличие исчисляется в лучшем случае 4.4% (в Западной Африке – Гвинея-Бисау, Гамбия, Сенегал, Мали, Нигер, Марокко), и 0.4% среди афроамериканцев. Этому же вторит статья Scozzari et al. (2012), что им удалось найти в Африке для геномных иследований только двоих (!) носителей A1a-M31. Ниже приведен список из 44 снипов гаплогруппы А1b, но в той системе, которую использует YFull. В классификации ISOGG в гаплогруппе A1b насчитывается 47 снипов, и ни один не пересекается по индексам с приведенными ниже. Из этих 47 снипов у меня есть L989, L1009, L1013, L1053, P108, V221. Остальных нет, потому что, как указывалось выше, не все снипы у каждого определяются. Список снипов гаплогруппы A1b (согласно классификации, используемой YFull)
Третья вилка – расхождение А1 на A1a и A1b. Потомки A1a сейчас живут в Африке и по всему миру (в частности, в ряде европейских стран и на Ближнем Востоке). Снипы А1b есть у всех нас, читатели Переформата. Но это опять не значит, что наши предки вышли из Африки. Это опять промежуточные снипы, и никого с терминальными снипами А1b пока не выявлено, ни в Африке, ни вне Африки. Поэтому называть его «африканским» опять никакого смысла не имеет. По данным, приведенным YFull, к A1a ведет 495 снипов (это 71 тысяча лет, если снип образуется в среднем раз в 144 года), к А1b – 44 снипа (6300 лет), но почему-то им опять было выставлено по 132600 лет «возраста». Опять загадка, на каком основании так делается. Хотя носители гаплогруппы A1a разбросаны по всему миру (поэтому в любом случае это не «африканская» гаплогруппа, даже по такому условному критерию, как современное место жительства), их мало. И.Р. Рожанский уже приводил в своей статье «Африканцы на севере Европы. Африканцы ли?», что гаплогруппа A1a-M31 чрезвычайно редкая, и даже в Африке ее наличие исчисляется в лучшем случае 4.4% (в Западной Африке – Гвинея-Бисау, Гамбия, Сенегал, Мали, Нигер, Марокко), и 0.4% среди афроамериканцев. Этому же вторит статья Scozzari et al. (2012), что им удалось найти в Африке для геномных иследований только двоих (!) носителей A1a-M31. Ниже приведен список из 44 снипов гаплогруппы А1b, но в той системе, которую использует YFull. В классификации ISOGG в гаплогруппе A1b насчитывается 47 снипов, и ни один не пересекается по индексам с приведенными ниже. Из этих 47 снипов у меня есть L989, L1009, L1013, L1053, P108, V221. Остальных нет, потому что, как указывалось выше, не все снипы у каждого определяются. Список снипов гаплогруппы A1b (согласно классификации, используемой YFull)  И вот здесь снипы гаплогруппы A1b приоткрывают загадку. Воспроизведем еще раз фрагмент лесенки снипов гаплогруппы A1:
И вот здесь снипы гаплогруппы A1b приоткрывают загадку. Воспроизведем еще раз фрагмент лесенки снипов гаплогруппы A1:  Мы видим, что от нее вилкой отходят субклады A1a и A1b. Мы уже узнали, что А1а встречается в Европе, на Ближнем Востоке, в Африке, причем нет никаких данных, что он изначально был африканским. Субклад A1b встречается у всех неафриканцев как промежуточный, в цепочке снипов. Субкладов, нижестоящих от A1b, а это A1b1, A1b1a, A1b1a1, A1b1a1a, A1b1a1a1, A1b1a1a2, A1b1a1a2a, A1b1a1a2b – у неафриканцев нет, возможно, потому, что параллельно с A1b1 образовался субклад BT (по даным YFull примерно 126 тысяч лет назад), и A1b1 ушли в Африку, а ВТ остался вне Африки. То есть у неафриканцев имется основная линия A1b > BT, и потомки ее составляют все гаплогруппы от В до Т. И вдруг выясняется, что нижестоящая линия, в субкладе A1b1a1- M23, P3, обнаружилась в Южной Африке среди носителей койсанских языков. Но это – довольно далекий по цепочке вниз субклад, который, по-видимому, отошел от неафриканцев в древние времена. Иначе говоря, не неафриканцы отошли от африканцев, а наоборот, африканцы отошли от неафриканцев, мигрировав в Африку. Опять – не выходили наши предки из Африки. Было наоборот: современные африканцы – потомки неафриканцев. Четвертая вилка – расхождение A1b на A1b1 и BT. A1b1 и нисходящие субклады считаются африканскими – по нынешнему месту проживания его носителей, хотя родительская гаплогруппа A1b (как промежуточный субклад в цепочке снипов) насчитывает не менее 2-3 миллиардов неафриканцев, а нижеследующие субклады от A1b1 и ниже – обнаружены всего у нескольких африканцев. Вот и считайте, кто от кого произошел. Ответ ясен – африканцы от неафриканцев. Повторю – не выходили наши предки из Африки. Итак, потомки A1b1 сейчас живут в минимальном количестве в Африке, а потомки гаплогруппы ВТ – это все неафриканские гаплогруппы от В до Т. По данным YFull, к A1b1 ведут всего 26 снипов, к субкладу A1b1a ведут всего 2 снипа, к A1b1a1 – 11 снипов, ни одного из них у нас нет. Датировок ни к одному из нисходящих от A1b1 субкладов в списке YFull нет, и понятно, почему – их носители крайне редко встречаются, да и данных нет. Снипы ВТ есть у всех нас, читатели Переформата. Список снипов гаплогруппы A1b1 (согласно классификации, используемой YFull)
Мы видим, что от нее вилкой отходят субклады A1a и A1b. Мы уже узнали, что А1а встречается в Европе, на Ближнем Востоке, в Африке, причем нет никаких данных, что он изначально был африканским. Субклад A1b встречается у всех неафриканцев как промежуточный, в цепочке снипов. Субкладов, нижестоящих от A1b, а это A1b1, A1b1a, A1b1a1, A1b1a1a, A1b1a1a1, A1b1a1a2, A1b1a1a2a, A1b1a1a2b – у неафриканцев нет, возможно, потому, что параллельно с A1b1 образовался субклад BT (по даным YFull примерно 126 тысяч лет назад), и A1b1 ушли в Африку, а ВТ остался вне Африки. То есть у неафриканцев имется основная линия A1b > BT, и потомки ее составляют все гаплогруппы от В до Т. И вдруг выясняется, что нижестоящая линия, в субкладе A1b1a1- M23, P3, обнаружилась в Южной Африке среди носителей койсанских языков. Но это – довольно далекий по цепочке вниз субклад, который, по-видимому, отошел от неафриканцев в древние времена. Иначе говоря, не неафриканцы отошли от африканцев, а наоборот, африканцы отошли от неафриканцев, мигрировав в Африку. Опять – не выходили наши предки из Африки. Было наоборот: современные африканцы – потомки неафриканцев. Четвертая вилка – расхождение A1b на A1b1 и BT. A1b1 и нисходящие субклады считаются африканскими – по нынешнему месту проживания его носителей, хотя родительская гаплогруппа A1b (как промежуточный субклад в цепочке снипов) насчитывает не менее 2-3 миллиардов неафриканцев, а нижеследующие субклады от A1b1 и ниже – обнаружены всего у нескольких африканцев. Вот и считайте, кто от кого произошел. Ответ ясен – африканцы от неафриканцев. Повторю – не выходили наши предки из Африки. Итак, потомки A1b1 сейчас живут в минимальном количестве в Африке, а потомки гаплогруппы ВТ – это все неафриканские гаплогруппы от В до Т. По данным YFull, к A1b1 ведут всего 26 снипов, к субкладу A1b1a ведут всего 2 снипа, к A1b1a1 – 11 снипов, ни одного из них у нас нет. Датировок ни к одному из нисходящих от A1b1 субкладов в списке YFull нет, и понятно, почему – их носители крайне редко встречаются, да и данных нет. Снипы ВТ есть у всех нас, читатели Переформата. Список снипов гаплогруппы A1b1 (согласно классификации, используемой YFull)  К сводной гаплогруппе ВТ ведут 403 снипа, и многие из них есть у нас. У гаплогрупп категории А их, естественно, нет. Датировка ВТ по данным YFull – примерно 126 тысяч лет, хотя 403 снипа – это примерно 58 тысяч лет. Некоторые могут подумать, что эти снипы – от родительской гаплогруппы A1b, но та датирована той же компанией YFull примерно 133 тысячами лет назад, то есть всего на 7000 лет раньше. Опять остается неясным, к чему те 403 снипа относятся, и откуда тянутся. Самая загадка в том, как «параллельные» гаплогруппы A1b1 и BT, от одного общего предка A1b, и к которым идут 26 и 403 снипа, соответственно, могут оказаться с одинаковой датировкой – 126 тысяч лет. Ниже приведен частичный список снипов сводной гаплогруппы ВТ. Как уже отмечалось, многие из них есть у каждого неафриканского мужчины. У меня есть, например, L418, L604, L962, L970, L977, PF1408 и другие. Фрагмент из списка снипов гаплогруппы BT (приведены 166 снипов из 403)
К сводной гаплогруппе ВТ ведут 403 снипа, и многие из них есть у нас. У гаплогрупп категории А их, естественно, нет. Датировка ВТ по данным YFull – примерно 126 тысяч лет, хотя 403 снипа – это примерно 58 тысяч лет. Некоторые могут подумать, что эти снипы – от родительской гаплогруппы A1b, но та датирована той же компанией YFull примерно 133 тысячами лет назад, то есть всего на 7000 лет раньше. Опять остается неясным, к чему те 403 снипа относятся, и откуда тянутся. Самая загадка в том, как «параллельные» гаплогруппы A1b1 и BT, от одного общего предка A1b, и к которым идут 26 и 403 снипа, соответственно, могут оказаться с одинаковой датировкой – 126 тысяч лет. Ниже приведен частичный список снипов сводной гаплогруппы ВТ. Как уже отмечалось, многие из них есть у каждого неафриканского мужчины. У меня есть, например, L418, L604, L962, L970, L977, PF1408 и другие. Фрагмент из списка снипов гаплогруппы BT (приведены 166 снипов из 403)  Итак, первая часть этой статьи безоговорочно показывает, что «африканские ДНК-линии» совсем не древние, в противовес тому, чем нас пичкают популяционные генетики уже 30 лет. Если вести «древность» от общих предков с приматами, то африканские линии ничем не отличаются от неафриканских, имеют одну и ту же глубину. Если считать от времени жизни общих предков современных мужских популяций – по племенам или по территориям, то африканские ДНК-линии весьма мелкие, намного мельче, чем неафриканские. Если считать от образования гаплогрупп, то никто не знает, на каких территориях или континентах они образовались, поэтому присваивать им названия «африканские» – некорректно. Более того, носителей этих гаплогрупп, категории А, в Африке очень мало. К этому могут быть разные причины – прошли бутылочные горлышки популяции, вымерли, или прибыли в Африку относительно недавно – большого значения не имеет, они не «древние», в том смысле, который вкладывают попгенетики. В итоге имеем то, что изображено на заглавной иллюстрации поста. Так что с этим разделом закончили – нет в Африке «древних ДНК-линий». Но есть такое понятие, как совокупные ДНК-линии. Это тогда, когда сами ДНК-линии мелкие, но они – ошметки древних раскидистых деревьев гаплогрупп и субкладов. Мы видим только верхушки, но экстраполяция верхушек, в данном случае гаплотипов этих «ошметок», позволяет увидеть, когда жили общие предки этих разбросанных гаплотипов. Мы уже видели это выше – сравнение гаплотипов «мелких» популяций гаплогрупп А00 и А0 показывает, что они расходятся на астрономические расстояния. Сами «ошметки» имеют общих предков всего несколько сотен лет назад, но они расходятся на полмилиона лет, и отсюда следует, что их общие предки жили около 250 тысяч лет назад. В этом отношении африканские ДНК-линии отличаются от неафриканских. Рассмотрим это подробнее, в этом упрятана одна из основных загадок человечества. Загадка была впервые сформулирована в 2012 году, в статье (Klyosov, Rozhanskii, Advances in Anthropology, 2, No. 2, 80-86). Суть ее состояла в том, что все ДНК-линии современных гаплогрупп от В до Т, то есть неафриканских гаплогрупп, сходятся к общему предку, который жил 64000±6000 лет назад, хотя разрозненные линии африканских (по нынешнему месту жительства) гаплогрупп сходятся к общему предку 132 тысяч лет назад (хотя сами разрозненные линии датируются временами жизни общих предков всего лишь 5500, 5000 и 600 лет назад). При этом все неафриканские линии (включая гаплогруппу В) отстоят исключительно далеко от африканских линий, ясно показывая, что неафриканские линии не могли произойти от африканских. Например, базовые гаплотипы гаплогрупп А0 и В отстоят друг от друга на 270 тысяч лет (между ними 18 мутаций в «медленных» 22-маркерных гаплотипах; для сравнения, базовые гаплотипы R1a и R1b отстоят друг от друга всего на 8 мутаций, то есть примерно на 54 тысячи лет, и это помещает их общего предка на 27 тысяч лет назад), иллюстрация увеличивается по клику.
Итак, первая часть этой статьи безоговорочно показывает, что «африканские ДНК-линии» совсем не древние, в противовес тому, чем нас пичкают популяционные генетики уже 30 лет. Если вести «древность» от общих предков с приматами, то африканские линии ничем не отличаются от неафриканских, имеют одну и ту же глубину. Если считать от времени жизни общих предков современных мужских популяций – по племенам или по территориям, то африканские ДНК-линии весьма мелкие, намного мельче, чем неафриканские. Если считать от образования гаплогрупп, то никто не знает, на каких территориях или континентах они образовались, поэтому присваивать им названия «африканские» – некорректно. Более того, носителей этих гаплогрупп, категории А, в Африке очень мало. К этому могут быть разные причины – прошли бутылочные горлышки популяции, вымерли, или прибыли в Африку относительно недавно – большого значения не имеет, они не «древние», в том смысле, который вкладывают попгенетики. В итоге имеем то, что изображено на заглавной иллюстрации поста. Так что с этим разделом закончили – нет в Африке «древних ДНК-линий». Но есть такое понятие, как совокупные ДНК-линии. Это тогда, когда сами ДНК-линии мелкие, но они – ошметки древних раскидистых деревьев гаплогрупп и субкладов. Мы видим только верхушки, но экстраполяция верхушек, в данном случае гаплотипов этих «ошметок», позволяет увидеть, когда жили общие предки этих разбросанных гаплотипов. Мы уже видели это выше – сравнение гаплотипов «мелких» популяций гаплогрупп А00 и А0 показывает, что они расходятся на астрономические расстояния. Сами «ошметки» имеют общих предков всего несколько сотен лет назад, но они расходятся на полмилиона лет, и отсюда следует, что их общие предки жили около 250 тысяч лет назад. В этом отношении африканские ДНК-линии отличаются от неафриканских. Рассмотрим это подробнее, в этом упрятана одна из основных загадок человечества. Загадка была впервые сформулирована в 2012 году, в статье (Klyosov, Rozhanskii, Advances in Anthropology, 2, No. 2, 80-86). Суть ее состояла в том, что все ДНК-линии современных гаплогрупп от В до Т, то есть неафриканских гаплогрупп, сходятся к общему предку, который жил 64000±6000 лет назад, хотя разрозненные линии африканских (по нынешнему месту жительства) гаплогрупп сходятся к общему предку 132 тысяч лет назад (хотя сами разрозненные линии датируются временами жизни общих предков всего лишь 5500, 5000 и 600 лет назад). При этом все неафриканские линии (включая гаплогруппу В) отстоят исключительно далеко от африканских линий, ясно показывая, что неафриканские линии не могли произойти от африканских. Например, базовые гаплотипы гаплогрупп А0 и В отстоят друг от друга на 270 тысяч лет (между ними 18 мутаций в «медленных» 22-маркерных гаплотипах; для сравнения, базовые гаплотипы R1a и R1b отстоят друг от друга всего на 8 мутаций, то есть примерно на 54 тысячи лет, и это помещает их общего предка на 27 тысяч лет назад), иллюстрация увеличивается по клику.  Ясно, что если между гаплогруппами А0 и В 270 тысяч лет, то вторая никак не могла образоваться от первой в течение всего периода существования Homo sapiens, не говоря о 70 тысячах лет назад, которые попгенетики отводят для времени образования гаплогруппы В из А. Это аналогично примеру выше для R1a и R1b, в котором 54 тысячи лет расстояния между ними никак не могут допустить, что одна образовалась из другой; они обе образовались от одного общего предка, которым в данном случае являлась гаплогруппа R1. Гаплогруппа А0, далеко отстоящая от гаплогруппы В, здесь не исключение. Подобные огромные расстояния отделяют гаплогруппу В от любых гаплотипов в субкладах категории А, например, 250 тысяч лет от A0a-L981, 280 тысяч лет от A1b-M32, 310 тысяч лет от европейского кластера гаплогруппы A1a-M31, 360 тысяч лет от другого кластера той же гаплогруппы, 320 тысяч лет от арабского кластера той же гаплогруппы, 350 тысяч лет от A0a-V150, 356 тысяч лет от A0a-L979, 346 тысяч лет от A0a1a-V151, и так далее. Самое большое расстояние отделяет гаплогруппу В от А00 – 450 тысяч лет, то есть их общий предок жил примерно 225 тысяч лет назад, на утренней заре Homo sapiens. Такая же картина наблюдается и для гаплотипов всех гаплогрупп от В до Т. Например, растояние от гаплогруппы С до А0-L896 составляет 357 тысяч лет, до A0a1-V150 – 300 тысяч лет, и так далее. Но расстояние от В до С составляет всего 120 тысяч лет, то есть их общий предок жил примерно 60 тысяч лет назад. Это находится в полном соответствии с картиной, что гаплогруппа В образует единый кластер с гаплогруппами от В до Т, и не является по происхождению «африканской» гаплогруппой. Видимо, уже всем ясно, что гаплогруппа В не образовалась из какой-либо гаплогруппы категории А (поскольку нет гаплогруппы А как таковой, есть А00, А0-Т, А0, А1, А1a, A1b и так далее), а все они ведут свое начало от общего предка. Этот общий предок жил примерно 160 тысяч лет назад, как показано в упомянутой выше статье (Klyosov, Rozhanskii, Advances in Anthropology, 2, No. 2, 80-86). Если исходить из датировок, приведенных на сайте YFull, то основатели гаплогрупп А00 и A0-T жили раньше этого времени, причем гаплогруппа А00 осталась в известном смысле тупиковой, нисходящие от нее субклады и прочие ДНК-линии неизвестны. A0-T прошла своими снипами как к африканцам, так и к неафриканцам, как показано выше, поскольку образовалась еще до разделения на африканскую и неафриканскую ветви. Разделение на эти ветви соответствует временам непосредственно до образования гаплогрупп А0 и А1, причем А0 ушла в Африку, А1 осталась вне Африки. Впоследствии носители многих гаплогрупп мигрировали в Африку, например, A1a и нижестоящие субклады, A1b1 и нижестоящие субклады, поэтому их снипов у нас нет, как описано выше. Все это описано в целом в упомянутой выше статье (Klyosov, Rozhanskii, Advances in Anthropology, 2012), которая, кстати, набрала рекордное количество просмотров (около 90 тысяч) и скачиваний (около 20 тысяч), но академическое сообщество предпочло ее не заметить. Слишком многое она меняет в представлениях. Основная иллюстрация в той статье следующая:
Ясно, что если между гаплогруппами А0 и В 270 тысяч лет, то вторая никак не могла образоваться от первой в течение всего периода существования Homo sapiens, не говоря о 70 тысячах лет назад, которые попгенетики отводят для времени образования гаплогруппы В из А. Это аналогично примеру выше для R1a и R1b, в котором 54 тысячи лет расстояния между ними никак не могут допустить, что одна образовалась из другой; они обе образовались от одного общего предка, которым в данном случае являлась гаплогруппа R1. Гаплогруппа А0, далеко отстоящая от гаплогруппы В, здесь не исключение. Подобные огромные расстояния отделяют гаплогруппу В от любых гаплотипов в субкладах категории А, например, 250 тысяч лет от A0a-L981, 280 тысяч лет от A1b-M32, 310 тысяч лет от европейского кластера гаплогруппы A1a-M31, 360 тысяч лет от другого кластера той же гаплогруппы, 320 тысяч лет от арабского кластера той же гаплогруппы, 350 тысяч лет от A0a-V150, 356 тысяч лет от A0a-L979, 346 тысяч лет от A0a1a-V151, и так далее. Самое большое расстояние отделяет гаплогруппу В от А00 – 450 тысяч лет, то есть их общий предок жил примерно 225 тысяч лет назад, на утренней заре Homo sapiens. Такая же картина наблюдается и для гаплотипов всех гаплогрупп от В до Т. Например, растояние от гаплогруппы С до А0-L896 составляет 357 тысяч лет, до A0a1-V150 – 300 тысяч лет, и так далее. Но расстояние от В до С составляет всего 120 тысяч лет, то есть их общий предок жил примерно 60 тысяч лет назад. Это находится в полном соответствии с картиной, что гаплогруппа В образует единый кластер с гаплогруппами от В до Т, и не является по происхождению «африканской» гаплогруппой. Видимо, уже всем ясно, что гаплогруппа В не образовалась из какой-либо гаплогруппы категории А (поскольку нет гаплогруппы А как таковой, есть А00, А0-Т, А0, А1, А1a, A1b и так далее), а все они ведут свое начало от общего предка. Этот общий предок жил примерно 160 тысяч лет назад, как показано в упомянутой выше статье (Klyosov, Rozhanskii, Advances in Anthropology, 2, No. 2, 80-86). Если исходить из датировок, приведенных на сайте YFull, то основатели гаплогрупп А00 и A0-T жили раньше этого времени, причем гаплогруппа А00 осталась в известном смысле тупиковой, нисходящие от нее субклады и прочие ДНК-линии неизвестны. A0-T прошла своими снипами как к африканцам, так и к неафриканцам, как показано выше, поскольку образовалась еще до разделения на африканскую и неафриканскую ветви. Разделение на эти ветви соответствует временам непосредственно до образования гаплогрупп А0 и А1, причем А0 ушла в Африку, А1 осталась вне Африки. Впоследствии носители многих гаплогрупп мигрировали в Африку, например, A1a и нижестоящие субклады, A1b1 и нижестоящие субклады, поэтому их снипов у нас нет, как описано выше. Все это описано в целом в упомянутой выше статье (Klyosov, Rozhanskii, Advances in Anthropology, 2012), которая, кстати, набрала рекордное количество просмотров (около 90 тысяч) и скачиваний (около 20 тысяч), но академическое сообщество предпочло ее не заметить. Слишком многое она меняет в представлениях. Основная иллюстрация в той статье следующая:  Номенклатура в этой иллюстрации несколько устаревшая, в ней нет гаплогруппы А00, которая в 2012 году была неизвестной, но суть ее остается правильной. Гаплогруппа альфа – это условная гаплогруппа, от которой пошло разделение эволюционного Y-хромосомного дерева человечества на африканскую (слева) и неафриканскую линии. В настоящее время все гаплогруппы от В до Т, то есть гаплогруппы неафриканского происхождения, сходятся к условной же бета-гаплогруппе, которой на самом деле является сводная гаплогруппа ВТ. Загадка – куда делась связка между альфа- и бета-гаплогруппой, протяженностью в сто тысяч лет? И, кстати, куда делись африканские ДНК-линии, которые на левой части диаграммы полностью экстраполированы, от общих предков всего от тысячи до нескольких тысяч лет давности, вглубь времен? Со времени публикации той статьи прошло уже более трех лет, в базах данных и в Проектах с тех пор накопилось много гаплотипов, в том числе протяженных, 111-маркерных, и появилась возможность проверить количественные выводы статьи. То, что расхождение африканской и неафриканской ДНК-линий произошло примерно 160 тысяч лет назад, мы уже проверили. Перейдем ко времени жизни общего предка всех неафриканских гаплогрупп – от В до Т. Ниже – дерево гаплотипов этих гаплогрупп. Оно построено по 111-маркерным гаплотипам, и сами гаплогруппы показаны по окружности дерева. Для каждой из 18 гаплогрупп использовали 10-25 гаплотипов, взятых произвольно из выборки, всего 403 гаплотипа nd 111-маркерном формате. Гаплогруппы категории А на дереве не оставили, так как они явно «вылезали» из кругового дерева, поскольку, как мы уже показали, они удалены по мутациям от всех других гаплогрупп.
Номенклатура в этой иллюстрации несколько устаревшая, в ней нет гаплогруппы А00, которая в 2012 году была неизвестной, но суть ее остается правильной. Гаплогруппа альфа – это условная гаплогруппа, от которой пошло разделение эволюционного Y-хромосомного дерева человечества на африканскую (слева) и неафриканскую линии. В настоящее время все гаплогруппы от В до Т, то есть гаплогруппы неафриканского происхождения, сходятся к условной же бета-гаплогруппе, которой на самом деле является сводная гаплогруппа ВТ. Загадка – куда делась связка между альфа- и бета-гаплогруппой, протяженностью в сто тысяч лет? И, кстати, куда делись африканские ДНК-линии, которые на левой части диаграммы полностью экстраполированы, от общих предков всего от тысячи до нескольких тысяч лет давности, вглубь времен? Со времени публикации той статьи прошло уже более трех лет, в базах данных и в Проектах с тех пор накопилось много гаплотипов, в том числе протяженных, 111-маркерных, и появилась возможность проверить количественные выводы статьи. То, что расхождение африканской и неафриканской ДНК-линий произошло примерно 160 тысяч лет назад, мы уже проверили. Перейдем ко времени жизни общего предка всех неафриканских гаплогрупп – от В до Т. Ниже – дерево гаплотипов этих гаплогрупп. Оно построено по 111-маркерным гаплотипам, и сами гаплогруппы показаны по окружности дерева. Для каждой из 18 гаплогрупп использовали 10-25 гаплотипов, взятых произвольно из выборки, всего 403 гаплотипа nd 111-маркерном формате. Гаплогруппы категории А на дереве не оставили, так как они явно «вылезали» из кругового дерева, поскольку, как мы уже показали, они удалены по мутациям от всех других гаплогрупп.  Видно, что все гаплогруппы от В до Т образуют достаточно единую систему ДНК-линий (выраженную здесь через 111-маркерные гаплотипы), так что про некую «африканскую гаплогруппу В» говорить не приходится. Она такая же «автохтонная африканская», как R1b – «автохтонная европейская». Это застарелая болезнь попгенетиков – то, что видят сейчас, значит, так было всегда. Болезнь называется «отсутствие научного подхода», или «отсутствие научной школы», что, впрочем, одно и то же, а если по-русски, то «думать не умеют». 403 гаплотипа в 111-маркерном формате, то есть почти 45 тысяч аллелей – это крепкий орешек, если считать вручную. Более того, за десятки тысяч лет в них накапливаются десятки тысяч мутаций, помимо того, огромное количество возвратных мутаций, что делает 111-маркерную систему такой древности неустойчивой, и неприемлемой для расчетов, поскольку датировки значительно занижаются. Поэтому для данных случаев разработана 22-маркерная панель из самых «медленных» маркеров. Они настолько устойчивы во времени, что для всех 403 гаплотипов за десятки тысяч лет маркер DYS472 вообще не мутировал ни разу, DYS436 – всего 30 раз, DYS590 – 42 раза, и так далее. Сравните – маркер DYS390 за это же время должен мутировать 2270 раз, то есть в среднем 5.6 раз на гаплотип. При таких частотах мутации расчет вклада обратных мутаций уже практически невозможен, система нестабильна. А вот «медленная» 22-маркерная панель с малыми скоростями мутаций позволяет проводить расчеты на десятки и сотни тысяч лет вглубь, и даже на миллионы лет, как было показано выше на примере расчета времени жизни общих предков человека и шимпанзе. Более того, разработанный калькулятор Килина-Клёсова проводит расчеты по каждому маркеру отдельно, используя квадратичный метод, и поэтому не требует введения поправок на возвратные мутации. В итоге ДНК-генеалогия получила незаменимый метод расчета массивных серий гаплотипов в любом формате и любой древности общего предка (Килин и Клёсов, Вестник Академии ДНК-генеалогии, 2015, т. 8, № 3, 321-375, http://dna-academy.ru/kilin-klyosov/). Расчет дерева из 403 гаплотипов, приведенного выше, показал, что общий предок гаплогрупп от В до Т жил примерно 63 тысячи лет назад (результат без округления – 63036 лет, что, конечно, с такой точностью совершенно излишне). Это почти точно совпадает с рассчитанной ранее (Klyosov and Rozhanskii, 2012) величиной 64±6 тысяч лет. Теперь осталось подойти к решению сформулированной в той статье загадки – повторим ее опять: куда же делись предки Y-хромосомных линий в Африке ранее 5-10 тысяч лет назад, и у неафриканцев ранее 64±6 тысяч лет назад? Причем у африканских линий группы А можно косвенным путем, а именно при сравнении протяженных гаплотипов с «неглубокими» общими предками, достичь времен жизни общих предков 132 тысячи лет назад, а у неафриканцев, как их гаплотипы ни комбинировать и ни сравнивать, глубже 64±6 тысяч лет уйти не получается. Более древних общих предков как корова языком слизнула. Что же это за корова была такая? Некоторые предположения в цитированной выше статье сделаны были – упомянуто возможное последствие извержения вулкана Тоба в Индонезии, около 70 тысяч лет назад. Не исключено было резкое похолодание, возможно, как результат извержения того же вулкана, причем, по мнению специалистов, ранг катастрофичности глобального похолодания намного выше, чем извержения вулкана, но это все оставалось довольно абстрактными предположениями. И вообще – чтобы извержение вулкана в Индонезии привело к гибели почти всех людей на планете – это кажется маловероятным, но все это «по понятиям». Нужно было мнение специалиста, пусть мнение и спорное, неоднозначное, но главное – подтвержденное конкретными данными. И такой специалист по данной проблеме есть. Знакомьтесь – Валерий Павлович Юрковец, профессиональный геофизик, палеоклиматолог, специалист в области влияния климатических катастроф, в том числе импактного характера, на миграции древнего человека, член Академии ДНК-генеалогии с начала ее образования несколько лет назад. Продолжение часть 2 Анатолий А. Клёсов, доктор химических наук, профессор Источник
Видно, что все гаплогруппы от В до Т образуют достаточно единую систему ДНК-линий (выраженную здесь через 111-маркерные гаплотипы), так что про некую «африканскую гаплогруппу В» говорить не приходится. Она такая же «автохтонная африканская», как R1b – «автохтонная европейская». Это застарелая болезнь попгенетиков – то, что видят сейчас, значит, так было всегда. Болезнь называется «отсутствие научного подхода», или «отсутствие научной школы», что, впрочем, одно и то же, а если по-русски, то «думать не умеют». 403 гаплотипа в 111-маркерном формате, то есть почти 45 тысяч аллелей – это крепкий орешек, если считать вручную. Более того, за десятки тысяч лет в них накапливаются десятки тысяч мутаций, помимо того, огромное количество возвратных мутаций, что делает 111-маркерную систему такой древности неустойчивой, и неприемлемой для расчетов, поскольку датировки значительно занижаются. Поэтому для данных случаев разработана 22-маркерная панель из самых «медленных» маркеров. Они настолько устойчивы во времени, что для всех 403 гаплотипов за десятки тысяч лет маркер DYS472 вообще не мутировал ни разу, DYS436 – всего 30 раз, DYS590 – 42 раза, и так далее. Сравните – маркер DYS390 за это же время должен мутировать 2270 раз, то есть в среднем 5.6 раз на гаплотип. При таких частотах мутации расчет вклада обратных мутаций уже практически невозможен, система нестабильна. А вот «медленная» 22-маркерная панель с малыми скоростями мутаций позволяет проводить расчеты на десятки и сотни тысяч лет вглубь, и даже на миллионы лет, как было показано выше на примере расчета времени жизни общих предков человека и шимпанзе. Более того, разработанный калькулятор Килина-Клёсова проводит расчеты по каждому маркеру отдельно, используя квадратичный метод, и поэтому не требует введения поправок на возвратные мутации. В итоге ДНК-генеалогия получила незаменимый метод расчета массивных серий гаплотипов в любом формате и любой древности общего предка (Килин и Клёсов, Вестник Академии ДНК-генеалогии, 2015, т. 8, № 3, 321-375, http://dna-academy.ru/kilin-klyosov/). Расчет дерева из 403 гаплотипов, приведенного выше, показал, что общий предок гаплогрупп от В до Т жил примерно 63 тысячи лет назад (результат без округления – 63036 лет, что, конечно, с такой точностью совершенно излишне). Это почти точно совпадает с рассчитанной ранее (Klyosov and Rozhanskii, 2012) величиной 64±6 тысяч лет. Теперь осталось подойти к решению сформулированной в той статье загадки – повторим ее опять: куда же делись предки Y-хромосомных линий в Африке ранее 5-10 тысяч лет назад, и у неафриканцев ранее 64±6 тысяч лет назад? Причем у африканских линий группы А можно косвенным путем, а именно при сравнении протяженных гаплотипов с «неглубокими» общими предками, достичь времен жизни общих предков 132 тысячи лет назад, а у неафриканцев, как их гаплотипы ни комбинировать и ни сравнивать, глубже 64±6 тысяч лет уйти не получается. Более древних общих предков как корова языком слизнула. Что же это за корова была такая? Некоторые предположения в цитированной выше статье сделаны были – упомянуто возможное последствие извержения вулкана Тоба в Индонезии, около 70 тысяч лет назад. Не исключено было резкое похолодание, возможно, как результат извержения того же вулкана, причем, по мнению специалистов, ранг катастрофичности глобального похолодания намного выше, чем извержения вулкана, но это все оставалось довольно абстрактными предположениями. И вообще – чтобы извержение вулкана в Индонезии привело к гибели почти всех людей на планете – это кажется маловероятным, но все это «по понятиям». Нужно было мнение специалиста, пусть мнение и спорное, неоднозначное, но главное – подтвержденное конкретными данными. И такой специалист по данной проблеме есть. Знакомьтесь – Валерий Павлович Юрковец, профессиональный геофизик, палеоклиматолог, специалист в области влияния климатических катастроф, в том числе импактного характера, на миграции древнего человека, член Академии ДНК-генеалогии с начала ее образования несколько лет назад. Продолжение часть 2 Анатолий А. Клёсов, доктор химических наук, профессор Источник Расширение сельского хозяйства с Ближнего Востока в Европу (9500-3800 до н.э.)
Расширение сельского хозяйства с Ближнего Востока в Европу (9500-3800 до н.э.) Распространение гаплогруппы J2 в Европе
Распространение гаплогруппы J2 в Европе Распределение гаплогруппы J1 в Европе, на Ближнем Востоке и в Северной Африке
Распределение гаплогруппы J1 в Европе, на Ближнем Востоке и в Северной Африке  Распределение гаплогрупп E1b1b в странах Европы, Ближнего Востока и Северной Африки
Распределение гаплогрупп E1b1b в странах Европы, Ближнего Востока и Северной Африки