Урок по теме: "История развития экологических связей человечества. Древние гоминиды". Древние гоминиды презентация
"История развития экологических связей человечества. Древние гоминиды" страница 3
Вывод: Прародиной человека считают Южную Африку, где были найдены останки приматов, названных парапитеками (латинский корень питек означает «обезьяна»). Эти останки датируют примерно 4 - 5 мил. лет
В настоящее время большинство ученых утверждают, что люди современного физического типа появились в Африке и оттуда мигрировали в другие районы, замещая там архаичные человеческие группы. Таким образом, вероятнее всего, прародиной древнейших гоминид и людей современного физического типа являлась Африка. ( слайд 14-15).
Дают определение слову антропогенез и отвечают на вопрос о теориях происхождения человека.
Доклад учащихся. Задают вопросы докладчикам
Работают с учебником. Поисковая беседа.
Слушают учителя.
2 мин
10-12 мин
2-3 мин
15 мин
Закрепление изученного материала
Антропогенез освещает проблемы
1) происхождения, эволюции человека
2) мобильности человека
3) эволюции человека
4) обитания на определенной территории
Прародина человека:
А) Австралия
Б) Южная Америка
В) Юго-Западная Европа
Г) Восточная Африка
Задание 2. Выберите три правильных утверждения из шести.
1. Специфические черты человека:
А) S-образный изгиб позвоночника Б) сводчатая стопа В) вторая сигнальная система Г) длинные передние конечности Д) развитая лицевая часть черепа Е) кисть с противопоставленным большим пальцем
2. Биологические факторы эволюции человека:
А) генотипическая изменчивость Б) естественный отбор В) коллективный образ жизни Г) трудовая деятельность Д) популяционные волны Е) членораздельная речь
Задание 3. Подберите соответствия.
1. Пути биологического прогресса, обеспечившие появление признаков человека:
«Гоминид» переводится как «человекоподобный», и в данное биологическое семейство входят, в том числе и люди. Вообще, в нём выделяют четыре основные ветви: люди, шимпанзе, гориллы и орангутаны. Ранее в семейство Гоминид включали лишь людей, в остальные три ветви объединяли в семейство Понгид. Однако, в современной биологии принято объединять их всех в большое семейство Гоминиды, уже в нём выделяя два подсемейства:
Понгины – сюда входят орангутаны.
Гоминины – сюда входят люди, шимпанзе и гориллы.
Однако, в антропологии сохранилась другая система, согласно которой к гоминидам относятся только люди и их вымершие предки. В данном случае, мы будем отталкиваться именно от антропологического определения, и проведём сравнительный анализ гоминидов, выделим их общие признаки, а также рассмотрим основные стадии эволюции.
Рис. 1 – Гоминиды
Общие признаки гоминидов
От остальных приматов гоминидов отличает целый ряд признаков, возникающих постепенно по мере антропогенеза. Современный человек имеет как сходства с человекообразными обезьянами, так и отличия от них. Первые обусловлены его происхождением, вторые же тем, что от жизни на деревьях он перешёл к обитанию на земле, в результате чего у него развилось прямохождение и трудовая деятельность – организм изменился, приспосабливаясь к новым условиям.
Выделим признаки адаптации гоминидов к передвижению на двух ногах:
туловище и шея расположены вертикально, при этом нижние конечности полностью разогнуты в коленном суставе;
тазобедренные мышцы развиты и расположены специфическим образом, форма таза расширена;
позвоночник изогнут, имеет S-образную форму;
грудная клетка уплощена;
череп уравновешен;
мышцы ноги сильно развиты;
стопа сводчатая, пальцы на ней укоротились, первый палец развит сильнее остальных и не противопоставлен им;
пропорции тела в целом сильно изменились.
Изменения строения руки гоминидов из-за её приспособления к труду:
произошло укорочение руки;
прочность кисти стала выше, как и её подвижность;
скелет большого и указательного пальца усилился;
большой палец удлинён;
мышца, противопоставляющая его, развилась сильнее, обособилась мышца, осуществляющая его сгибание;
развилась способность к ротации руки;
сильнее развились отделы мозга, отвечающие за движения кисти.
Изменения в строении мозга гоминида:
мозг увеличился в размерах;
кора и ассоциативные зоны сильно развились;
также развились теменная и лобная доли, разрослись и развились их цитоархитектонические поля;
получили развитие речевые центры;
межнейрональные контакты усилились;
мозолистое тело стало крупнее;
борозды третьего порядка стали многочисленнее.
Также произошла редукция волосяного покрова, он поредел и укоротился, изменилось строение гортани из-за усиления речевой функции – произошло развитие хрящей и связок, изменилась топография гортани.
Специфические признаки черепа и зубов, отличающие гоминидов:
лицевой отдел черепа при сравнении с мозговым уменьшился;
нижняя челюсть стала меньше;
клыки уменьшились;
гребни на черепе исчезли;
носовой отдел стал выступать;
подбородочный выступ развился;
профиль лица в целом выпрямился.
Основные виды гоминидов
Австралопитековые. Находки их многочисленны, в результате чего скелет известен целиком. Обитали в большей части Африки, за исключением северной части. Самые древние гоминиды – наиболее ранние австралопитековые (рис. 2), жили примерно 7 миллионов лет назад. Выделяют два основных рода: Australopithecus, также называемые грацильными австралопитеками, и Paranthropus или массивные австралопитеки.
Скелет говорит о том, что состоялось прямохождение, длина тела: у грацильных – 100-130 см., у массивных – 150-178 см. Рука освободилась от опорной функции, но ещё сохраняла многие примитивные особенности, строение её нестабильно. Большой палец ещё не полностью противопоставлен остальным, фаланги изогнуты, кроме конечных, которые плоские.
Объём мозга в среднем составлял 450 кубических сантиметров для грацильных особей, и немногим больше 500 куб. см. – для массивных. Клыки и резцы малых размеров, коренные зубы крупные. Челюсти более массивны, чем у человека. Имели выступающее надбровье, при небольшом размере лица в целом, срезанную носовую область и закруглённый затылочный отдел. Имели выраженный половой диморфизм: размеры самцов превышали размеры самок.
Рис. 2 – Австралопитековые гоминиды
Величина и степень изношенности коренных зубов говорит о том, что питались австралопитеки твёрдой пищей, например, злаками. Если массивные австралопитеки являлись исключительно растительноядными, то грацильные были всеядными.
Исходя из числа находок, считается, что общая численность австралопитеков составляла примерно от 10 до 20 тысяч. Средняя продолжительность жизни составляла 19,7 лет. Являясь общественными гоминидами, жили они стадами, каждое из которых состояло из нескольких десятков особей, включая нескольких, взрослых самцов, а также большее число самок и детёнышей. Это позволяло при необходимости противостоять хищникам. Маловероятно, что существовала элементарная семья, поскольку ей препятствовала необходимость групповой охоты, а также первоначальная трудовая деятельность, требовавшая участия десятков особей.
Архантропы (рис. 3). Ареал обитания – Евразия и Африка. Обитали в Африке примерно за 1,6 млн. лет до нашего времени, чуть позже появились в Азии, и лишь 500 тысяч лет назад – в Европе. Тело длиннее, чем у австралопитеков, достигая, для мужских особей, 175 см. Кости длинные и толстые, скелет массивен. Череп имеет толстые стенки, затылочный и надглазничный валики мощные, убегающая лобная часть, крупные челюсти. Крупными были также и зубы. Рельеф затылочного отдела сильно развит, поскольку для балансировки головы, при прямохождении, требовалась мощная мускулатура.
Средний объём мозга превышал 1000 куб. м. Лобные доли развиты не так сильно, как у современного человека, височные и теменные доли узкие. Зоны неокортекса подверглись усиленному развитию из-за развития речевых функций и ручного труда. Усиливается рельеф мышц языка, начинает оформляться подбородок.
Рис. 3 – Архантроп
Общая численность оценивается по-разному: от десяти десятков тысяч до миллиона особей. Группы архантропов имели большую численность, чем у австралопитеков, составляя в среднем около полусотни особей. В сообществах часто происходили конфликты, также отмечены случаи каннибализма, которые трактуются по-разному – либо как убийство ради еды, либо как поедание убитого в результате конфликта из-за самок.
Детская смертность составляла 40%, из-за чего средняя продолжительность жизни едва ли превышала таковую у австралопитеков, однако, если вычесть умерших до 14 лет, то она составит 38 лет.
Гейдельбергский человек. Их стали выделять с относительно недавних пор, ранее же относили либо к архантропам, либо к палеоантропам. Однако, они имеют немало особенностей, позволивших выделить их в отдельную эволюционную стадию. Время обитания – 800-200 тысяч лет назад. Произошли от архантропов. Данный вид политипичен, имеет широкий ареал распространения – от самого юга Африки, и до севера континентальной Европы (исключая Скандинавию).
Мозговая полость меньше, чем у неандертальцев, а высота свода черепа больше, лоб менее покат, скуловые кости не скошены, мозговой отдел имеет меньшую длину, орбиты относительно невысокие, сосцевидные отростки развились.
В Европе поначалу обитали на юге, затем начали перемещаться к северу, всё более приобретая черты неандертальцев. Гейдельбергский человек (рис. 4) занимался изготовлением орудий из дерева, кости, камня, делал постройки, занимался изготовлением одежды. Отмечено, что гейдельбержцы старались придать своим орудиям более эстетичный вид. В остатках поселений имеются постоянные очаги и семейные постройки.
Рис. 4 – Гейдельбергский человек
Палеоантропы (рис. 5). Обитали примерно в промежутке 300-30 тысяч лет до нашей эры в Евразии и Африке. Поздние неандертальцы выделяются небольшим ростом (до 170 см. у мужских особей), шириной плеч, развитием скелета и мускулатуры. Пропорции тела схожи с нынешними представителями народов Арктики в результате адаптации к погодным условиям. Массивное строение скелета принято объяснять приспособлением к необходимости переносить много мяса на большие расстояния.
Объём мозга в среднем 1350 куб. см., отмечается рост неокортекса. Череп крупных размеров, форма удлинена, толщина стенок превышает таковую у современных людей. Надглазничные валики массивные, лоб покат, челюсти большие и массивные, а зубы крупные.
Рис. 5 – Палеонтроп
Есть теории, утверждающие, что онтогенез скелета у неандертальцев происходил быстрее, чем у современных людей. При этом проводятся сравнения останков неполовозрелых неандертальцев с современными детьми. Эти теории отмечают быстрый темп взросления как важную особенность неандертальцев.
Численность неандертальцев среднего палеолита оценивается в миллион. Увеличения продолжительности жизни в сравнении с предыдущими формами не произошло, редкостью был неандерталец, доживший до 30 лет, средняя же продолжительность жизни составляла около 20 лет.
Имели сложную социальную организацию, демонстрировали дружелюбное социальное поведение, а также альтруизм, выражавшийся, в частности, в заботе о старых и больных особях. Социальные роли были дифференцированы, уровень кооперации высокий. Однако, практиковался каннибализм, хотя с течением времени всё в меньшей степени. В это время появились первые погребения. Согласно данным археологии, общины неандертальцев были склонны замыкаться, в результате чего начиналось родственное скрещивание и происходили биологические изменения. Возможно, с этим связана потеря ими эволюционной пластичности и последовавшее вымирание данного вида.
Ископаемые люди современного типа. Кроманьонцы современного типа полностью сложились примерно 40 тысяч лет назад. Отличия от современного человека невелики. В первую очередь – массивный костяк, большая толщина костей черепа и больший размер глазниц, меньшая высота свода.
Общая численность составляла примерно от 3 до 5,5 миллионов, в том числе примерно 50 тысяч в Европе. Средняя продолжительность жизни для взрослого – 34 года, более трети переживало сорокалетний рубеж. Возникла дуально-общинная организация. Так как она несла преимущества в области генетики, размеры групп возросли.
vse-lekcii.ru
"История развития экологических связей человечества. Древние гоминиды" страница 2
Мы рассмотрели историю развития жизни на земле с момента ее возникновения до настоящего времени. Так какая наука изучает происхождение человека?
Антропогенезом –
называют процесс выделения человека из мира животных. (слайд 6).
Существует множество теорий происхождения человека. Какие из них Вам знакомы? Ответ:
1) Панспермия
2) Креационизм
3) Эволюционная теория или теория Дарвина. (слайд 7).
Научные гипотезы о происхождении человека. (слайд 8-10).
Физминутка для улучшения мозгового кровообращения( слайд 11).
Учитель: Гипотезы о прародине человека.
Но где находилась прародина человека? В каком районе нашей планеты протекали самые ранние стадии антропогенеза?
Палеонтологи, антропологи и археологи в качестве возможных центров зарождения человечества называли самые различные районы Земли. Наиболее часто — Африку и Южную Азию. Это объясняется тем, что именно в этих районах были обнаружены останки рамапитеков, имевших прогрессивные признаки.
Накопленные в настоящее время научные данные позволяют считать "колыбелью человечества" Северо-Восточную Африку. Хотя некоторые исследователи до сих пор отстаивают Южную Азию, но в пользу африканской гипотезы говорят такие факты как:
В Африке в настоящее время живут антропоморфные обезьяны, из всех приматов наиболее близкие к человеку.
В Южной Африке обнаружены остатки существ, которые по всей вероятности, были двуногими и по строению передних зубов и всего скелета в целом обладали исключительно большим сходством с человеком.
Диффференциации высших приматов, выработка у них разных типов локомоции и превращение каких-то групп двуногих наземных антропоморфных в древнейших людей должна была благоприятствовать очень большая величина африканского континента и разнообразие его ланшафтов.
Данные о телантропе позволяют предполагать, что в Южной Африке в начале плейстоцена существовали формы, еще более близкие человеку, чем австралопитеки.
Новейшие открытия показали, что находки типа рамапитека, в Африке древнеее, чем в Сиваликских холмах, кроме того, такие формы как Homo habilis и чадантроп – являются более прогрессивными формами, чем австралопитеки. (слайд 12).
Но в настоящее время большинство ученых считают, что судьба процесса гоминизации оказалась разной на азиатском и африканском континентах, В плиоцене (примерно 7—8 млн лет назад) в Южной Азии резко изменился климат. Условия жизни стали неблагоприятными для гоминид, что привело их к вымиранию или переселению в другие районы земного шара.
В Африке, особенно в ее восточной приэкваториальной части, наоборот, сложились очень благоприятные условия для жизни гоминид. Поэтому здесь процесс гоминизации продолжался, и гоминиды достигли такого уровня, когда они уже полностью отделились от животных и приобрели специфические человеческие черты. (слайд 13).
В конце XX в. свой вклад в решение проблемы происхождения человека стали вносить молекулярная биология и генетика. Для выяснения времени и места появления человека современного типа стали использовать молекулярно-генетические методы. Так, анализ мирового распределения изменчивости структуры митохондриальной ДНК, проведенный американскими исследователями, позволил им предположить, что люди современного типа появились южнее Сахары около 200 тыс. лет назад. Ученые установили, что численность популяции в период видообразования составляла около 10 000 человек. В настоящее время большинство ученых считают, что люди современного физического типа появились в Африке и оттуда мигрировали в другие районы, замещая там архаичные человеческие группы . . (слайд 13)
infourok.ru
Эволюция и климатические изменения
Важнейшим фактором эволюции были глобальные климатические изменения, которые вели к изменениям вмещающего ландшафта и перестройки всей среды обитания. В эволюции предков человека, большая часть которой протекала на территории Африки, такие изменения сыграли очень большую роль. Период с 23 до 1,7 мил. лет назад характеризуется постепенным похолоданием и иссушением (аридизации). В следствии этого площади тропических лесов, в которых обитал гоминоиды, а затем и гоминиды, постепенно сокращались, преобладавшие ранее закрытые ландшафты (джунгли) замещались открытыми (саванны). Сезонные колебания температуры и влажности становились все более резкими.
Наиболее ощутимые изменения приходились на время 8–7 мил. лет назад. Резкое сокращение площади тропических лесов привело к значительному сокращению пищевых ресурсов и обострили конкуренцию между различными видами, в том числе и обезьян. В этой борьбе явно выигрывали мелкие древесные формы. Так число видов гоминоидов сократилось в несколько раз, а число видов собаковидных в тот же время увеличилось в несколько раз.
В этих условиях единственным выходом для гоминоидов (человекообразных обезьян) был поиск и освоение новых экологических ниш, недоступных конкурентам. В результате они стали осваивать нижние ярусы леса и примыкавшие к джунглям саванны. Примерно в тоже время в Африке происходили тектонические изменения – возникла Восточноафриканская рифтовая система – гигантский разлом с окаймляющими его горными хребтами. Это привело к тому, что климат в Восточной Африке стал сухим, а оставшиеся здесь гоминоиды были изолированы от других районов и были вынуждены интенсивно приспосабливаться к быстро меняющимся условиям жизни.
Эволюция семейства гоминид.
Так в период ок. 8 – 4,5 мил. лет назад началось выделение семейства гоминид из гоминоидов.
1. Древнейший гоминид
Середина 90-х гг. стало переломным временем – резкое увеличение находок позволило ученым изменить и уточнить наше представление о древнейшей истории гоминид.
Заметим, что ранее (70-е гг.) в качестве исходного рода гоминидной линии рассматривалирамапитеков(жили от 15 до 7 мил. лет назад), сейчас считается, что они стоят у иной линии, ведущей к орангутангам, а не людям. В настоящее время с сточки зрения систематики этот исходный вид еще не выделен. Существующие гипотезы (например, уранопитек) оспариваются. Постоянно поступают новые данные (B Сахаре нашли древний череп неизвестного человекоподобного существа).
2. Первые известные гоминиды
2.0. На протяжении многих лет ранним (но не первым) видом гоминидов более или менее хорошо известны австралопитеки Австралопитек в Википедии.
Но раскопки последних лет дали новый материал, который позволил сделать предположение о существовании более ранних видов. Кратко расскажем о них.
2.1. В 2000 г. на западе Кении были найдены фрагменты бедренной и плечевой костей, которые позволили выделить новый вид гоминид – оррорин тугененсис, который существовал ок. 6 мил. лет назад. Чуть позже, в 2001–2002 гг. был найден еще более ранний вид, который получил название сахельантроп. Благодаря выделению этих видов стало ясно, что гоминидная линия выделилась не древнее, чем 5 мил. лет назад. Заметим, что с этим согласны не все ученые.
2.2. Ранее в 1994 г. на севере Эфиопии был открыт самый ранний вид гоминид, который получил название Ардипитек рамидус – сначала его считали самым древним австралопитеком, но потом выделили в особой отдельный вид и род. Сейчас известно несколько видов рамидусов – которые жили в период от 5,8 до 4,4 мил. лет назад.
Причины выделения новой линии связывают с резким изменением климатических условий (похолодание ок. 5 мил лет назад).
2.3. В настоящее время выделено несколько видов австралопитеков и прослежена эволюция этого вида на протяжении нескольких миллионов лет с 4,2 по 1 мил. лет назад.
А
встралопитеки жили от 4 до 1 мил. лет назад. До 3 мил. лет назад они расселялись только в восточной Африки, после этого расселились в центре и на юге это континента.
так выглядел череп австралопитека (использован материал сайта Библиотека "Ан Ома Ли Я?" – электронная библиотека посвященная необъяснимым явлениям, загадкам Мировой и Русской истории...)
Они обладали прямохождением (см. Найденный в южноафриканских пещерах австралопитек ходил почти вертикально) – анатомия таза близка к человеку. Они имели сводчатую стопу, непротивопоставляемый большой палец стопы и т.п. Этим они отличались от всех остальных гоминид. Рост 100 – 160 см. Объем мозга от 400 до 500 куб. см. Морфология австралопитеков поражает сочетанием прогрессивного по строению скелета и примитивного черепа.
Самый древний вид австралопитеков – австралопитек анаменсис (4,2 – 3,9 мил лет назад), далее австралопитек афарский (3,5 мил. назад). Заметим, что на основании находок 1998 – 1999 г. ученые выяснили, что параллельно с австралопитеками афарским ок. 3,5 лет назад сосуществовал еще один род гоминид – кениантроп. После этого возникло, как минимум, еще 7 видов. Все они разделяются на «грацильных» (ав-к афарский, ав-к африканский, ав-к гархи) и «массивные» (зинджантроп бойсов, парантроп робустус, парантроп эфиопский). Массивные возникли позже. Но и исчезли позже (ок. 1 мил. лет назад). Самым массивным австралопитеком был зинджантроп. Находки датируются 1,8 мил. лет назад. Объем черепа ок. 530 см. куб. (см. подробности, детали)
Кто из них стал предшественником вида Хомо – в настоящее время науке неизвестно !
Австралопитеки немалую часть времени еще проводили на деревьях, здесь они находили и еду и надежной убежище и спокойный ночлег. Возможно, были стоянки и на земле.
Формы организации: бродячая группа, всеядны. Продолжительность жизни ок. 17–22 лет. Австралопитеки могли использовать некоторые природные предметы в качестве орудий добывания пищи. Австралопитеки вымерли ок. 1 мил. лет назад.
3. В 3 – 2,5 тыс. до н.э. существование гоминид было под угрозой. Начался цикл иссушения климата, ландшафты Африки изменились, сократились осадки и стало холоднее. Усилилась необходимость приспосабливаться к новым условиям.
Следующим этапом антропогенеза стало появления вида Хомо хабилис (человек умелый). Древнейшие представители вида хомо известны ок. 2,4 – 2,3 мил. лет назад. Название обусловлено нахождением орудий труда. Рядом с останками этого вида гоминид находят первые орудия труда, изготовленные преднамеренно. Ученые считаю, что неизвестно, происходят ли они от одной из форм австралопитеков или от гоминид какого-либо другого вида. Начинается первый самый ранний период истории.
Это интересно
studfiles.net
Презентация на тему "Предки человека"
Презентация на тему: Предки человека
Скачать эту презентацию
Скачать эту презентацию
№ слайда 1 Описание слайда:
Предки человека
№ слайда 2 Описание слайда: № слайда 3 Описание слайда: № слайда 4 Описание слайда:
Дриопитек ДРИОПИТЕКИ (Dryopithecus, т. е. древесная обезьяна) — единственный род вымерших человекообразных обезьян, подсемейства Dryopithecinae: включает три подрода (собственно дриопитеки, сивапитеки и проконсулы) и несколько видов. Остатки дриопитеков (нижние челюсти, зубы, кости конечностей) известны из миоценовых и плиоценовых отложений Западной Европы, Восточной Африки и Южной Азии (Индия). ДРИОПИТЕКИ (Dryopithecus, т. е. древесная обезьяна) — единственный род вымерших человекообразных обезьян, подсемейства Dryopithecinae: включает три подрода (собственно дриопитеки, сивапитеки и проконсулы) и несколько видов. Остатки дриопитеков (нижние челюсти, зубы, кости конечностей) известны из миоценовых и плиоценовых отложений Западной Европы, Восточной Африки и Южной Азии (Индия).
№ слайда 5 Описание слайда:
Особенности дриопитеков При передвижении по деревьям эти обезьяны цеплялись за ветви, подвешиваясь к ним руками, туловище при этом находилось в отвесном положении, а ноги были поджаты. У дриопитековых были особенности, позволявшие им идти по пути антропогенеза: высокое развитие центральной нервной системы, хорошее цветное бинокулярное зрение и хватательные конечности – не только передние, но и задние. Способность к передвижению на двух ногах, или к прямохождению, очень пригодилась предкам человека, когда на Земле стал сильно меняться климат.Великий русский физиолог И. Павлов выделял обезьян среди др. животных. Благодаря 4 хватательным конечностям у обезьян складываются более разнообразные взаимоотношения с окружающей средой. Это, в свою очередь, развивает мышечное чувство, осязание, зрение: обезьяны видят предметы в объёме и цвете.
№ слайда 6 Описание слайда:
Рамапитек Сегодня многие ученые склоняются к мысли, что наиболее вероятным предком существ, развивающихся в направлении человека, является рамапитек. Останки его были найдены в Индии. (Находку назвали "рамапитек" в честь индийского бога Рамы: Рама и "питекос" - греческое слово - означает "обезьяна"). Это, по сути, точка, в которой пути эволюции человека и высших обезьян разошлись. Рамапитек, по-видимому, жил в редком лесу и некоторую часть времени еще проводил на деревьях.
№ слайда 7 Описание слайда:
АВСТРАЛОПИТЕК Австралопитеки (от латинского australis — южный и греческого pithеkos — обезьяна) группа ископаемых высших приматов, кости которых впервые были обнаружены в пустыне Калахари (Южная Африка) в 1924, а затем в Восточной и Центральной Африке.
№ слайда 8 Описание слайда:
Жизнь австралопитеков Австралопитеки населяли большую часть Африки в конце неогена в промежутке между 6 и 1 млн. лет назад. Они жили в саваннах семейными группами, включавшими 2–3 взрослых самцов, несколько самок и детенышей разных возрастов. Питались австралопитеки преимущественно растительной пищей, включая семена злаков, в связи с чем их клыки заметно уменьшились в размерах и перестали выступать над уровнем прочих зубов. Передвигались они на двух ногах, почти или полностью выпрямившись. Их мозг по объему лишь незначительно уступал мозгу наиболее древних людей рода Homo. Наиболее развитым он был у мелких видов австралопитеков. Африканский австралопитек (Australopithecus africanus) был ростом около 150 см, и, вероятно, пользовался примитивными орудиями — грубо оббитыми камнями, палками, обломками кости.
№ слайда 9 Описание слайда:
Пропорции тела Пропорции конечностей и морфология стопы и руки указывают на то, что существо из штеркфонтайнских пещер лазило по деревьям (используя мощный большой палец руки для тискообразного захвата), и передвигалось по земле на двух ногах. Таким образом, большой палец развился у древних предков людей именно для того, чтобы хвататься за ветки. У их потомков, относящихся к Homo, обитавших преимущественно на земле, он использовался уже для создания орудий труда и манипуляций.
№ слайда 10 Описание слайда:
Австралопитек Бойса Этот наиболее крупный и сильный из всех австралопитеков получил свое название в честь английского бизнесмена Чарльза Бойса, помогавшего финансировать палеонтологические исследования в Восточной Африке, в результате которых в 1959 г. был найден неизвестный ранее науке вид "южных обезьян". Крупный австралопитек Бойса показан рядом с современным человеком. Рост 1,6-1,78 м, но мог быть и меньше. Масса 60-80 кг.
№ слайда 11 Описание слайда:
Зубная дуга На этой неполной зубной дуге заметны признаки, характерные для вида Australopithecus boisei.а - Маленькие резцы, предназначенные для откусывания.б - Огромные коренные зубы, способные размалывать пищу.
№ слайда 12 Описание слайда:
Четыре изображения черепов A. boisei 1.Череп самца Australopithecus boisei. а - Сагиттальный стреловидный гребень. б - Реконструкция нижней челюсти. 2.Череп, принадлежавший, вероятно, самке, которая была значительно меньших размеров. 3. Боковой вид черепа самца. в - Надглазничный валик. 4. Вид черепа сверху. г - Скуловые дуги, к которым прикреплялись массивные челюстные мышцы
№ слайда 13 Описание слайда:
Австралопитек Южной Африки Stw 573 В 1978 г. был обнаружен наиболее древний австралопитек Южной Африки Stw 573, получивший также имя «Маленькая Нога» («Little Foot»). Находка представляет собой четыре сочлененных кости левой стопы от лодыжки до второго сустава большого пальца. Изначально останки были определены как принадлежащие мартышке, но в 1994 г., при пересмотре коллекций идентифицированы как кости австралопитека. Большой палец стопы сильно отстоял от остальных, обладал значительной подвижностью и, возможно, был хватательный; стопа Stw 573, таким образом, была одинаково приспособлена для лазания по деревьям.
№ слайда 14 Описание слайда:
Люси В 1974 году во впадине Афар Джохансон обнаружил частично сохранившийся скелет австралопитека, каталогизированный как AL 288-1, но более известный как «Люси», с оценочным возрастом 3,4 миллиона лет. Люси погибла во время какого-то катастрофического события – тело было изуродовано так, что даже кости таза оказались раздробленными еще во время гибели или спустя самое незначительное время; труп обезьяны как будто перемалывало в какой-то каменной мясорубке. Останки были найдены вмурованными в скальную породу на высоте 30 метров над нынешним уровнем реки Аваш, что говорило о значительном изменении рельефа местности с той поры или чересчур бурном потоке какого-то иного водного источника. Находка Люси явилась двойной удачей – впервые были найдены относительно полные (40 %) скелетные останки австралопитека, да еще столь древний возраст находки позволял присвоить ей звание нового основателя человеческого рода.
№ слайда 15 Описание слайда:
Люси При жизни Люси имела рост чуть более метра, вес чуть более 30 кг, и объем мозга в пределах сугубо обезьяньего – 400-420 см3. Ни о какой орудийной деятельности, разумеется, речи тоже идти не могло. Фрагмент тазовой кости Люси, восстановленный Оуэном Лавджоем из сорока разрозненных частей
№ слайда 16 Описание слайда:
На фото 1 – вскрытая археологами пещера Лонггупо в восточной китайской провинции Сычуань недавно преподнесла исследователям сенсационные находки – человеческие окаменелости и каменные экспонаты, возраст которых составляет около 2 миллионов лет по официальной шкале. Останки людей и их инструменты находятся в несомненной геологической ассоциации со многими окаменелостями млекопитающих того периода.
№ слайда 17 Описание слайда:
На фото 2 – фрагмент человеческой челюсти из китайской пещеры Лонггупо, который датируется возрастом 1,9 миллионов лет, с отдельно найденными резцами, характерными для Homo erectus/ergaster.
№ слайда 18 Описание слайда:
На фото 3 в верхней части – фрагмент человеческой челюсти из китайской пещеры Лонггупо (тот же, что и на фото 2), который очень напоминает аналогичный экземпляр с восточного берега озера Туркана (ER 992), датированный по официальной шкале приблизительно 1,2 млн. лет.
№ слайда 19 Описание слайда:
На фото 4 – каменные инструменты древних обитателей китайской пещеры Лонггупо. Образец А – каменный инструмент в стадии начальной обработки. Следы множества ударов, направленных в одну точку, исключают естественный характер повреждения образца. Артефакт В – один из самых ранних инструментов, известных в истории человечества – каменное рубило из китайской пещеры Лонггупо возрастом 1,9 млн. лет по официальной шкале. «Ненормальность» этой находки состоит в том, что при возрасте около 2 млн. лет она обнаружена не в Африке, а в Азии.
№ слайда 20 Описание слайда:
На фото 5 – обломок челюсти гоминида, близкого к Homo ergaster из Юаньмоу (Yuanmou), провинция Юнань (Yunnan), датируемый возрастом 5-6 млн. лет (!) по стандартной шкале.
№ слайда 21 Описание слайда:
На фото 6 – череп детеныша австралопитека, напоминающий знаменитого Бэби из Таунга (Australopithecus africanus). Обнаружен (в числе других австралопитековых фрагментов) в 1989 году в Юаньмоу, провинция Юнань, профессором Zhang Xingyong. Находка датируется средним плиоценом, то есть, приблизительно 4-5 млн. лет, что по возрасту превосходит аналогичные африканские экземпляры.
№ слайда 22 Описание слайда:
Строение черепа
ppt4web.ru
Происхождение и эволюция человека. Презентация. Основные этапы, стадии
Профессор кафедры антропологии биологического факультета МГУ, д.б.н.
От Редактора: перед Вами - текст доклада, прозвучавшего 24 февраля 2011 года на биологическом факультете МГУ.
Как правило, люди, далекие от антропологии, представляют себе теорию о происхождении человека консервативной и мало подверженной переменам. На самом деле современная эволюционная антропология – наиболее динамичная и изменчивая часть науки о человеке. Она во многом отличается от традиционного антропогенеза, с которым знакомили нас наши преподаватели еще, казалось бы, совсем недавно – в 60-е – 70-е годы прошлого века.
При этом концептуальный скелет эволюционной антропологии остается неизменным. Это представление о длительной биологической эволюции человека от древних приматов через целый ряд ископаемых гоминид к современному виду Homo sapiens. Однако частные концепции, формирующие тело, теоретическую плоть современной науки о происхождении человека, стремительно меняются. Так, на протяжении большей части 20-го века главной объяснительной теорией о причинах и факторах антропогенеза была так называемая трудовая теория Ф. Энгельса, согласно которой развитие прямохождения и головного мозга было напрямую связано с трудовой деятельностью и развитием руки. Ядро теории Ф. Энгельса составляла триада «рука – мозг – прямохождение», в которой обозначалась последовательность развития основных морфо-функциональных систем человеческой эволюции.
Далее, по мере накопления сведений о древнейших гоминидах трудовая теория антропогенеза Ф. Энгельса постепенно лишалась фактологической основы, а знаменитая триада «рука – мозг – прямохождение» утрачивала свою монолитность. Решающую роль в этом процессе сыграли находки австралопитеков и признание их эволюционной значимости. Согласно находкам, в то время как прямохождение уже стало привычным способом передвижения древнейших гоминид, их мозг по объему не превосходил мозг шимпанзе, а способность руки к трудовым операциям вызывала очень большие сомнения. В итоге к концу 20-го века только признак прямохождения остался главным и несомненным критерием гоминизации, то есть принадлежности ископаемых находок к семейству гоминид. Дискуссии в этой области велись лишь по поводу деталей – когда, почему и как возникло прямохождение, но сам способ передвижения сомнению не подвергался.
Но вот появляются новые находки, более подробно изучаются старые, и ситуация вновь меняется, неожиданно возникает новая проблема древней гоминидной локомоции вообще.
Вначале обнаружилось, что практически все австралопитековые, и даже ранние Homo обладали довольно странной особенностью, которую западные исследователи назвали «функциональной дихотомией», хотя более точно было бы обозначить её как «функциональная двойственность» [Johanson, Edey, 1981; Leakey et al., 1995; Alemseged, 2006]. Проявляется эта особенность в несколько разной степени выраженности комплекса прямохождения для верхних и нижних конечностей древнейших гоминид. Нижние конечности по толщине кортикального слоя длинных костей, форме и длине шейки бедра, расположению вертелов бедренной кости и другим особенностям полностью соответствуют прямохождению. В то же время строение верхних конечностей - пропорции, морфология костей плечевого пояса, изогнутость фаланг пальцев – говорит о способности к древесному передвижению. Как писала Е.Н. Хрисанфова, это можно рассматривать как свидетельство «широкого локомоторного репертуара» древнейших гоминид в условиях флуктуации климата и других факторов внешней среды [Хрисанфова, Перевозчиков, 1999].
Возможно и другое объяснение «функциональной дихотомии». Признаки древесного передвижения могли оказаться нейтральными и несколько «задержаться» на верхних конечностях, не мешая их владельцам практиковать полноценную бипедию.
Слайд 1. Ардипитек рамидус
Казалось бы, вопрос решен, объяснение найдено, и основные положения о значимости прямохождения как древней ключевой адаптации не пострадали. Однако новые находки показали, что не только верхние конечности древнейших гоминид были приспособлены к древолазанию, но и стопы некоторых из них имели явно обезьянье строение! Речь, прежде всего, идёт об ардипитеке с датировкой 4,4 млн. лет, находке которого был посвящен весь журнал «Science» в октябре 2009 г.
В этих публикациях ардипитек реконструируется как гоминид, который вел преимущественно древесный образ жизни, медленно перемещаясь по ветвям деревьев с опорой на стопы и ладони. Что касается наземного двуногого передвижения, то оно, по мнению исследователей, носило «эпизодический характер» [White et al., 2009]. Несмотря на приданное ардипитеку выпрямленное положение тела, нельзя не обратить внимание на довольно узкие тазовые кости, которые формируют таз явно понгидного типа. Но самое поразительное – это стопа ардипитека с резко противопоставленным большим пальцем! Согласно графику, приведенному в одной из статей, противопоставление большого пальца стопы у этого гоминида даже больше, чем у шимпанзе. Очень трудно представить себе развитую бипедию у примата с подобной конструкцией стопы! Авторы описания выстраивают гипотетическую эволюционную последовательность от ардипитека к австралопитеку афарскому, но в таком случае приходится допустить, что всего за 500 тыс. лет крайне специализированная стопа ардипитека могла эволюционировать в стопу афарского австралопитека с ее сводчатостью и приведенным первым пальцем. Слайд 1.
От редакции: рекомендуем ознакомиться с мнением нашего научного редактора на эту тему.
Слайд 2. Стопа Homo habilis OH 8
И здесь уместно вспомнить находку OH8, представляющую собой почти полную стопу, которую приписывают представителю самых ранних людей, Homo habilis. Наряду с чисто человеческими особенностями - укороченные пальцы, увеличенный большой палец, расположение пальцев в одной плоскости, полное развитие двойного свода стопы, - отмечаются следы хорошего развития задней большеберцовой мышцы, которая поворачивает стопу наружу, что может рассматриваться как маркер древолазания.
Возможно, в отношении древних гоминид вскоре можно будет говорить не только о широком локомоторном репертуаре, но о довольно широком спектре локомоторных адаптаций, представленных разными находками австралопитековых и даже ранних Homo. И в этом случае вновь возникает проблема критерия гоминизации – если это не рука, не крупный мозг и не устойчивое прямохождение, то что же? Получается, что могли успешно существовать частично древесные и четвероногие гоминиды? Могли ли такие существа быть предками рода Homo?
Очевидно, на все эти вопросы мы пока не можем дать ответ, но сама постановка подобных проблем, которые ещё недавно было просто невозможно себе представить, является знаковым признаком современной эволюционной антропологии.
В начале и середине 20-го века сформировалась и стадиальная теория антропогенеза, постулирующая последовательную смену эволюционных стадий, представленных конкретными находками. Согласно стадиальной теории, каждый эволюционный уровень представлен лишь одним вариантом гоминид, поэтому схемы эволюции неизбежно носят линейный характер, имеют вид ломаной линии, а каждая стадия отображается точкой на этой линии. Кроме того, предполагается, что гоминиды каждого эволюционного уровня являются носителями вполне определенной, доступной на их биологическом уровне культуры, отраженной в археологических орудиях. Согласно этим представлениям, развитие культуры идёт вслед за биологической эволюцией и зависит от неё.
Слайд 3.
Однако на сегодняшний день каждая эволюционная стадия оказывается представленной несколькими ископаемыми формами. Слайд 3. В отношении всех эволюционных этапов установлен факт высокого разнообразия одновременно существующих гоминидных вариантов. При этом в большинстве случаев мы ничего не можем сказать о том, как они соотносятся друг с другом. Исходя из данных о низком физиологическом барьере среди приматов в целом, а также из сложности критерия вида в отношении ископаемых форм, мы вправе предполагать некую возможность биологических смешений между этими формами. В этом случае наиболее приемлемой оказывается модель сетевидной, нелинейной эволюции, которая предполагает возможность генетических обменов между синхронными популяциями гоминид. Принимая концепцию сетевидной эволюции, мы вынуждены отказаться от традиционного представления об эволюционных линиях и в качестве представителей каждой эволюционной стадии рассматривать несколько вариантов гоминид. Идея сетевидной эволюции в современной антропологии наиболее ярко представлена гипотезой мультирегионального происхождения современного человека. Согласно мультирегиональной концепции, для эволюции человечества в планетарном масштабе оказалось вполне достаточно древнейшего исхода архантропов с территории африканского материка, после чего общность базового генофонда человечества поддерживалась благодаря сети локальных генных потоков.
Нелинейность эволюции можно признавать на словах, но принять это представление как основу наших взглядов на эволюцию человека оказывается весьма непросто. Проявлением линейного подхода в современной антропологии является модель «Out of Africa», в которой мы вновь имеем дело с эволюционными линиями. Африканская гипотеза предполагает последовательный исход с территории африканского континента нескольких миграционных волн, древнейшая из которых представлена архантропами, а самая поздняя – ранними сапиенсами.
Срабатывает инерция мышления, и одной из главных проблем для многих современных исследователей оказывается вопрос о том, какой же из одновременно существовавших вариантов гоминид дал начало дальнейшей прогрессивной линии эволюции (всё-таки линии!). С другой стороны, оперировать с понятием «эволюционная линия» намного более удобно, чем с понятием «пласт», и, выстраивая линейные схемы эволюции, многие современные исследователи подразумевают наличие в узловых точках неких обширных популяционных объединений.
Нетрудно видеть, что в модели мультирегиональной эволюции постоянные генные потоки между синхронными популяциями неизбежно формируют сменяющие друг друга пласты форм.
Слайд 4. Синхронные пласты гоминид и «эволюционный конус»
Автором в своё время был предложен термин «гетерогенный эволюционный пласт гоминид» для обозначения совокупности синхронных форм, репродуктивная изоляция которых не может быть ни доказана, ни опровергнута [Бахолдина, 1988]. Прогрессивные признаки могли быть рассеяны в пределах такого синхронного пласта, концентрируясь постепенно под действием отбора, по мере усиления процессов интеграции. Схема последовательно сменяющих друг друга эволюционных пластов может быть представлена в виде своеобразного конуса, внутри которого пунктиром обозначена магистральная линия прогресса, вокруг которой и происходит постепенная концентрация прогрессивных особенностей. За пределами конуса, когда видовой эволюционный полиморфизм гоминид сменяется видовым единообразием на уровне Homo sapiens sapiens, пунктир переходит в сплошную линию. Слайд 4.
Возникает вопрос – что дальше? Представляет ли эволюция современного человека линейный процесс? Скорее всего, нет, но эволюционные прогнозы - занятие неблагодарное. Необходимо добавить, что одна из наиболее плодотворных идей современной эволюционной антропологии, идея магистральной линии прогресса, подробно разработана в трудах А.А. Зубова [1985; 1996; 2004].
При этом нельзя сказать, что стадиальная теория эволюции полностью стала достоянием прошлого, ибо никто не отказывается от представления об эволюционных стадиях и их смене. Просто эти представления видоизменились и усложнились, вобрав в себя огромный массив новых данных.
Слайд 5.
Одной из особенностей нынешней ситуации в эволюционной антропологии является постоянное перемещение некоторых эволюционных пластов и отдельных находок «вверх» или «вниз» вдоль эволюционной вертикали. Относительно недавно, например, произошел дрейф единичной находки нижней челюсти, найденной в начале 20-го века недалеко от города Гейдельберг от уровня древнейших людей, архантропов, к более прогрессивному эволюционному пласту, в котором эта находка объединилась вместе с находками так называемых пренеандертальцев типа Сванскомб, Эрингсдорф и Штейнгейм. Образовался новый таксон, который стал называться, как и нижняя челюсть, Homo heidelbergensis, гейдельбергские люди. Сейчас этот таксон представлен обширным пластом находок из Африки, Азии и Европы. Другие находки попали в этот же таксон из более «высоких» пластов, как, например, знаменитый череп родезийского человека, который в учебниках антропологии 20-го века описывался в качестве африканского неандертальца. Слайд 5.
Представления о неандертальцах претерпели, пожалуй, наиболее радикальные изменения Главное из них состоит в перемещении неандертальцев от одного эволюционного уровня к другому – от уровня пусть спорной, но все же предковой формы, к уровню, занимаемому современным Homo sapiens. Слайд 5.
Сейчас неандертальцы и ископаемые сапиенсы рассматриваются как полиморфное человечество эпохи позднего плейстоцена (позже 200 тыс. лет).
Слайд 6. Находки, свидетельствующие о метисации между неандертальцами и сапиенсами
Антропологи пытаются решить вопрос: насколько это древнее человечество было единым? Иными словами, можно ли с уверенностью судить о наличии или отсутствии биологических смешений между сапиенсами и неандертальцами? Что касается ископаемых свидетельств, то существует довольно много находок, которые можно рассматривать в качестве морфологических подтверждений смешений между сапиенсами и неандертальцами. Это и известные находки с территории Передней Азии – Схул, Табун, Кафзех, Амуд, Кебара, и хорватское местонахождение Крапина, недавние находки с территории Испании и Португалии [Duarte et al., 1999; Schoenherr, 2008]. Слайд 6.
Однако до самого последнего времени генетики активно выступали против возможных смешений сапиенсов и неандертальцев, поскольку результаты изучения древней ДНК показывали очень большой масштаб различий между ними. Правда, постепенно накапливались и единичные генетические данные иного рода. Так, например, американские генетики показали, что самый распространенный вариант гена microcephalin, который определяет рост головного мозга, появился в популяции Homo sapiens 37 000 лет назад в результате скрещивания с каким-то другим архаичным видом. Этот вариант гена настолько сильно отличается от других, более редких его вариантов, которые встречаются у современных людей, что он не мог возникнуть в результате мутаций, а был получен извне. Появились и сведения о том, что связанный с речью ген FOXP2, с мутациями, характерными для сапиенсов, был обнаружен в ДНК неандертальцев.
Но самый сенсационный результат палеогенетических исследований был опубликован в мае прошлого года. Группа генетиков под руководством Сванте Паабо из Института эволюционной антропологии имени Макса Планка в Лейпциге расшифровала, наконец, 2/3 генома неандертальца. В качестве материала послужили кости скелета трех неандертальских женщин из пещеры в Хорватии. Оказалось, что генетические различия между неандертальцами и сапиенсами составляют всего 0,16 %! Гены, по которым наблюдаются различия, кодируют некоторые белки кожи, в том числе её пигментацию, связаны с энергетическим обменом, определяют степень активности мужских половых клеток.
Сравнение с отдельными группами современного человечества показало наибольшую близость неандертальцев к населению Евразии, и, прежде всего, к европейцам. Как пишет Елена Наймарк, излагая итоги этого исследования, «… среди древних вариантов аллелей имеются такие, которые существенно чаще встречаются у европейцев, чем у африканцев. Всего было известно 13 таких географически капризных вариантов. Так вот выяснилось, что 10 из этих 13 вариантов имеются и у неандертальцев!» http://elementy.ru/news/431316
Слайд 7. Происхождение, миграции и зоны смешений неандертальцев и сапиенсов Европы и Передней Азии
Результаты сравнения генома неандертальцев с современными обитателями разных регионов Земли представляются особенно интересными, потому что это тот редкий случай, когда данные генетики совпадают с данными палеоантропологии. Согласно нашим современным представлениям, неандертальцы – это древнейшее, коренное население Европы. Предполагается, что неандертальцы возникли в результате узконаправленной эволюции европейских популяций гейдельбергского человека. Из Европы отдельные группы неандертальцев могли мигрировать на юго-восток и расселиться на территории современного Кавказа и Передней Азии. Возможно, в этом регионе встретились миграционные потоки европейских неандертальцев и древнейших сапиенсов – выходцев из Африки. Слайд 7. Именно с территории Передней Азии и Европы известны находки смешанного типа, имеющие промежуточный между неандертальцами и сапиенсами морфологический облик.
Таким образом, мы теперь точно знаем, что смешения между двумя вариантами древнего человечества, действительно, имели место, и в наших жилах течет и неандертальская кровь! Но тогда неандертальцы и сапиенсы представляли собой не два разных вида, а две большие расы древнего человечества, и контакты между ними следует рассматривать не как межвидовые, а как межрасовые взаимоотношения. И в этом случае современные сапиенсы в некоторой степени оказываются потомками метисов между двумя верхнеплейстоценовыми расами человечества, что является еще одним ударом для сторонников «расовой чистоты», к которой, очевидно, наши предки были полностью равнодушны.
По-прежнему актуальной остается и проблема факторов антропогенеза. Многочисленные исследования в этой сфере обнаруживают высокую поливариантность и множественность причин и движущих сил человеческой эволюции. Большое значение, например, придается стратегиям питания, перемены в которых могли сыграть решающую роль в поворотные моменты биологической истории человечества.
Слайд 8.
Очень интересны исследования, касающиеся значимости полового отбора и фактора восприятия внешности, который сыграл важнейшую роль в исчезновении многих архаических и появлении современных признаков. Обычно роль этого фактора восстанавливается по результатам исследований о предпочтениях того или иного типа внешности современными мужчинами и женщинами. Однако последствия действия этого фактора можно обнаружить и на краниологическом материале. Автором было показано, что признаки лицевого скелета вносят наибольший вклад в общую морфологическую изменчивость и намного более эффективны при разграничении краниологических серий по сравнению с признаками мозгового черепа. Слайд 8. Возможно, эти различия носят эволюционный характер и сформировались в процессе становления вида Homo sapiens. На ранних этапах эволюции происходило ускоренное изменение размеров и формы мозгового черепа, а на поздних этапах началось интенсивное преобразование лицевого скелета под влиянием, в том числе, полового отбора, и антропологическая дифференциация человеческих популяций во многом происходила именно по этой системе признаков.
Меняются и основные тренды развития эволюционной антропологии. В начале и середине 20-го века главным вектором её движения был поиск новых находок, новых фактов и создание основных теоретических концепций, объясняющих эти факты. Сейчас одной из постоянных проблем является приведение в систему всего огромного разнообразия имеющихся данных. И в этом я вижу не просто сложную задачу, но некую ответственную миссию специалистов-антропологов, призванных не только выстраивать такую систему для самих себя, но и приобщать к ней широкий круг людей, заинтересованных в познании сложных процессов развития и становления современного человечества.
Литература:
Бахолдина В.Ю. Гетерогенный пласт гоминид как объект общего эволюционного процесса // Вопр. Антропологии. 1988. Вып. 81. С. 18 – 28.
Зубов А.А. Иерархия факторов антропогенеза // Вестник антропологии.1996. Вып. 2. С. 195 – 206.
Зубов А.А. Магистрилизация и демагистрализация в ходе эволюционного процесса // Вопр. Антропологии. 1985. Вып. 75. С. 14 – 26.
Зубов А.А. Палеоантропологическая родословная человека. М., 2004. 551 с.
Хрисанфова Е.Н., Перевозчиков И.В. Антропология. М.: Изд-во Московского университета, 1999. 400 с.
Alemseged Z., Spoor F., Kimbel W.H., Bobe R., Geraads D., Reed D., Wynn J.G. A juvenile early hominin skeleton from Dikika, Ethiopia // Nature. 2006. 443 (7109). P. 296–301.
Duarte C, Maurício J, Pettitt PB, Souto P, Trinkaus E, van der Plicht H, Zilhão J. The early Upper Paleolithic human skeleton from the Abrigo do Lagar Velho (Portugal) and modern human emergence in Iberia // Proceedings of the National Academy of Sciences 1999. 96 (13). P. 7604–7609.
Johanson, Donald; Maitland Edey (1981). Lucy: The Beginnings of Humankind. New York: Simon and Schuster, 1981. 416 р.
Leakey M. G., Feibel C. S., I. MacDougall & A. Walker. New four-million-year-old hominid species from Kanapoi and Allia Bay, Kenya // Nature. 1995. 376 (6541). P. 565–571.
Schoenherr N. Late Neandertals and modern human contact in southeastern Iberia // Washington University in St. Louis News & Information. 2008, 10 Dec. P. 1.
White T., Asfaw B., Beyene Y., Haile-Selassie Y., Lovejoy C.O., Suwa G., WoldeGabriel G. Ardipithecus ramidus and the Paleobiology of Early Hominids // Science. 2009. 326. P. 75 - 86.






 B Сахаре нашли древний череп неизвестного человекоподобного существа).
B Сахаре нашли древний череп неизвестного человекоподобного существа). Австралопитек в Википедии.
Австралопитек в Википедии. 

 Найденный в южноафриканских пещерах австралопитек ходил почти вертикально) – анатомия таза близка к человеку. Они имели сводчатую стопу, непротивопоставляемый большой палец стопы и т.п. Этим они отличались от всех остальных гоминид. Рост 100 – 160 см. Объем мозга от 400 до 500 куб. см. Морфология австралопитеков поражает сочетанием прогрессивного по строению скелета и примитивного черепа.
Найденный в южноафриканских пещерах австралопитек ходил почти вертикально) – анатомия таза близка к человеку. Они имели сводчатую стопу, непротивопоставляемый большой палец стопы и т.п. Этим они отличались от всех остальных гоминид. Рост 100 – 160 см. Объем мозга от 400 до 500 куб. см. Морфология австралопитеков поражает сочетанием прогрессивного по строению скелета и примитивного черепа.  подробности,
подробности,  детали)
детали) Описание слайда:
Описание слайда: 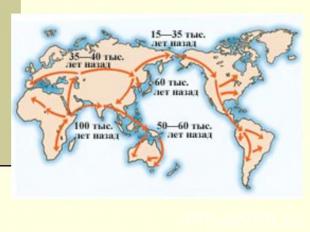 Описание слайда: № слайда 3
Описание слайда: № слайда 3 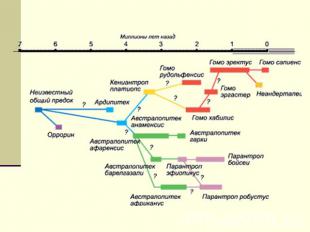 Описание слайда: № слайда 4
Описание слайда: № слайда 4  — единственный род") Описание слайда:
Описание слайда:  Описание слайда:
Описание слайда:  Описание слайда:
Описание слайда:  Описание слайда:
Описание слайда:  Описание слайда:
Описание слайда:  Описание слайда:
Описание слайда:  Описание слайда:
Описание слайда:  Описание слайда:
Описание слайда:  Описание слайда:
Описание слайда:  Описание слайда:
Описание слайда:  Описание слайда:
Описание слайда:  Описание слайда:
Описание слайда:  Описание слайда:
Описание слайда:  Описание слайда:
Описание слайда:  Описание слайда:
Описание слайда:  Описание слайда:
Описание слайда:  Описание слайда:
Описание слайда:  Описание слайда:
Описание слайда:  Описание слайда:
Описание слайда:  Профессор кафедры антропологии биологического факультета МГУ, д.б.н.
Профессор кафедры антропологии биологического факультета МГУ, д.б.н. 

















